LA Course #GCE-6-1501
About Tree Physiology
Edited by Len Phillips, updated January 2023
Sections Go directly to the section by clicking on the title below
About Tree Physiology
Edited by Len Phillips, updated January 2023
Sections Go directly to the section by clicking on the title below
Tree Physiology is the study of how trees grow and develop in terms of genetics, biochemistry, cells, tissues, and organ functions. It also relates to a tree’s interaction with environmental factors. While many physiological processes are similar in trees as in other plants, trees possess a unique physiology that help determine their outward appearance.
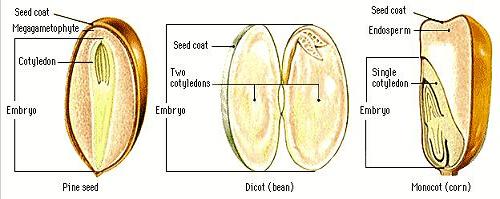
Have you ever wondered how trees begin? You know it is from seeds, but what tells the seed to begin to grow? Then you have to wonder, how does the seed know the conditions are just right so the embryo will be able to grow into a seedling and then into a tree? What parts of a tree begin to grow first?
Click on green text in each section for more information and photos.
Reproduction and Germination
Germination
Let's start with germination which is defined as the growth of an embryonic tree contained within a seed. It results in the formation of a tree seedling. Seed germination depends on both internal and external conditions being just right to support seedling growth. Often this depends on the individual seed variety and is closely linked to the ecological conditions of a tree's natural habitat.
When seeds are being formed, most trees store food as a reserve within the seed itself. The food consists of starch, proteins, and oils. This food reserve provides nourishment to push the growth of an embryonic tree. When the seed imbibes water, hydrologic enzymes are activated which break down these stored food resources into metabolically useful chemicals that will begin the growth process. There will be enough food to keep the embryonic tree growing until the primary leaves and roots begin the process of photosynthesis. The embryonic tree then becomes a seedling growing on its own.
Inducing Germination
The most important external factors to induce germination include temperature, water, oxygen, and sometimes light or darkness. Various trees require different environmental trigger levels for successful seed germination.
Germination
Let's start with germination which is defined as the growth of an embryonic tree contained within a seed. It results in the formation of a tree seedling. Seed germination depends on both internal and external conditions being just right to support seedling growth. Often this depends on the individual seed variety and is closely linked to the ecological conditions of a tree's natural habitat.
When seeds are being formed, most trees store food as a reserve within the seed itself. The food consists of starch, proteins, and oils. This food reserve provides nourishment to push the growth of an embryonic tree. When the seed imbibes water, hydrologic enzymes are activated which break down these stored food resources into metabolically useful chemicals that will begin the growth process. There will be enough food to keep the embryonic tree growing until the primary leaves and roots begin the process of photosynthesis. The embryonic tree then becomes a seedling growing on its own.
Inducing Germination
The most important external factors to induce germination include temperature, water, oxygen, and sometimes light or darkness. Various trees require different environmental trigger levels for successful seed germination.
- Temperature affects cell metabolism and growth rates. Seeds from different species and even seeds from the same tree germinate over a wide range of temperatures. This temperature range limits when they will germinate and they will not do so above or below the preferred range.
- Water is required for germination. Most seeds need enough water to moisten the seeds but not enough to soak them. The uptake of water by seeds is called imbibition, which leads to the swelling of the food reserves within the seed, which causes the breaking of the seed coat. Mature seeds are often extremely dry and need to take in significant amounts of water, relative to the dry weight of the seed before cellular metabolism and growth can begin.
- Oxygen is required by the germinating seed for metabolism. Oxygen is used in aerobic respiration, the main source of the seedling's energy until it grows leaves. Oxygen is found in pore spaces between the particles of soil. If a seed is buried too deeply within the soil or the soil is compacted or waterlogged, the seed can be oxygen starved and germination will be delayed. The seed will also die from rot caused by oxygen deprivation if too much water occurs after the germination process has started.
- Light or darkness can be an environmental trigger for germination and is a type of physiological dormancy. Most seeds are not affected by light or darkness, but many seeds, including species found in forest settings, will not germinate until an opening in the canopy allows sufficient light for growth of the seedling.
Seed Germination
All tree species can reproduce by seeds. Each species has a unique set of requirements for seed production and germination. Seed dispersal strategies vary widely, from wind-driven seeds such as on a maple (Acer sp.) tree, to seeds falling on the ground beneath the trees such as a nut tree, or to seeds carried away by certain species of animals or birds such as a crabapple being eaten by a bird and the seeds are dropped some distance from the tree.
Scarification
Any process of breaking, scratching, or altering the seed coat through mechanical, chemical, or thermal methods, such as freezing over winter, may be necessary to make the seed coat permeable to water and oxygen. This process is known as scarification. Some seeds have impermeable seed coats that prevent oxygen and water from entering the seed, causing a type of physical dormancy which is broken when the seed coat is worn away enough to allow gas exchange and water uptake from the environment. In nature, some seeds require particular conditions to germinate, such as the heat of a wildfire or soaking in a body of water for a long period of time. Others need to be passed through a bird or animal's digestive tract to weaken the seed coat enough to allow the embryonic tree to emerge.
For mechanical scarification, seed coats can be filed with a metal file, rubbed with sandpaper, nicked with a knife, or cracked gently with a hammer to weaken the seed coat. Scarification of seeds can also be achieved by imbibing them in concentrated sulfuric acid at appropriate concentrations and durations of treatment. Another scarification method involves the use of hot water for brief periods.
Breaking Dormancy
Some live seeds are dormant and need more time, and/or need to be subjected to specific environmental conditions before they will germinate. A good example is when a wildfire heats a cone and ejects the seed ready to germinate without competition from the burned forest trees. Seed dormancy can originate in different parts of the seed. Dormancy breaking often involves changes in membranes, initiated by certain hormones, notably abscisic acid, which inhibits germination, and gibberellin, which ends seed dormancy.
Roots
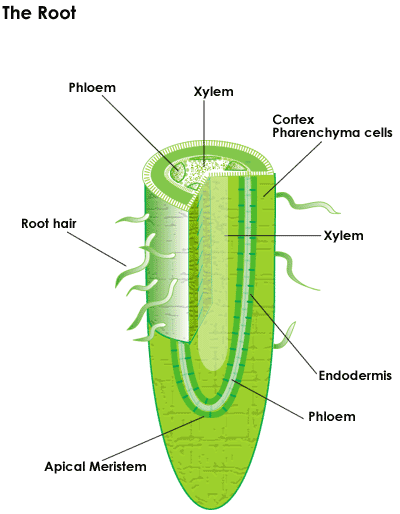
First to emerge from the seed coat is the primary root, or radicle as it is called in the embryo. This radicle is composed of one major root that is thicker at its base and tapers toward the growing tip. Early root growth is one of the functions of the apical meristem located near the tip of the root. The meristem cells more or less continuously divide, producing more meristem, root cap cells (these are sacrificed to protect the meristem), and undifferentiated root cells. The latter become the primary tissues of the root, first undergoing elongation. Gradually these cells differentiate and mature into specialized cells of root tissues.
After the embryonic seedling emerges from the seed coat and starts growing roots and leaves, the embryonic seedling's food reserves are quickly exhausted. At this point photosynthesis provides the energy needed for continuing growth and the seedling now requires a continuous supply of water, nutrients, and light. Most tree species grow a taproot from the seed until moisture or an obstacle is reached and the seedling can then focus on spreading structural roots, absorbing water and nutrients, and letting the leaves manufacture food.
Tree roots will generally grow in any direction where the correct environment of air, mineral nutrients, and water exists to meet the tree's needs. Roots will stop growing toward dry or poor soil conditions.
Vegetative Reproduction
Besides seeds, trees can be propagated with the assistance of mankind.
* Cultivars are propagated by grafting a bud or stem of the parent tree onto a seedling rootstock of the species.
* Clones are propagated by encouraging roots to develop on cuttings of the parent tree.
Sprouts and suckers start with dormant buds that "come alive" to form new shoots off the parent trees. Sprouts are shoots from stumps of a tree. Suckers are shoots that originate from buds on the root systems. Both sprouts and suckers have the potential to grow into new full size trees genetically identical to the parent. Often times, sprouts and suckers will not grow until the parent tree dies or becomes very sick or suffers from a severe injury. The buds are held in dormancy by hormones produced in the leaves. When these hormone levels drops below a certain point because of increased light level as the old tree thins its branches and leaves, the dormant buds begin to grow.
Vegetative layering is an uncommon form of vegetative propagation. When branches or stems come in contact with the soil, the cambium tissue sometimes will form roots. In this way, former branches might become trunks of several "new" trees genetically identical to the parent.
Tissue culture is an invention of modern science to extract cells of desirable trees and grow these cells into trees within laboratory conditions. Trees from this process are genetically identical to the parent.
All tree species can reproduce by seeds. Each species has a unique set of requirements for seed production and germination. Seed dispersal strategies vary widely, from wind-driven seeds such as on a maple (Acer sp.) tree, to seeds falling on the ground beneath the trees such as a nut tree, or to seeds carried away by certain species of animals or birds such as a crabapple being eaten by a bird and the seeds are dropped some distance from the tree.
Scarification
Any process of breaking, scratching, or altering the seed coat through mechanical, chemical, or thermal methods, such as freezing over winter, may be necessary to make the seed coat permeable to water and oxygen. This process is known as scarification. Some seeds have impermeable seed coats that prevent oxygen and water from entering the seed, causing a type of physical dormancy which is broken when the seed coat is worn away enough to allow gas exchange and water uptake from the environment. In nature, some seeds require particular conditions to germinate, such as the heat of a wildfire or soaking in a body of water for a long period of time. Others need to be passed through a bird or animal's digestive tract to weaken the seed coat enough to allow the embryonic tree to emerge.
For mechanical scarification, seed coats can be filed with a metal file, rubbed with sandpaper, nicked with a knife, or cracked gently with a hammer to weaken the seed coat. Scarification of seeds can also be achieved by imbibing them in concentrated sulfuric acid at appropriate concentrations and durations of treatment. Another scarification method involves the use of hot water for brief periods.
Breaking Dormancy
Some live seeds are dormant and need more time, and/or need to be subjected to specific environmental conditions before they will germinate. A good example is when a wildfire heats a cone and ejects the seed ready to germinate without competition from the burned forest trees. Seed dormancy can originate in different parts of the seed. Dormancy breaking often involves changes in membranes, initiated by certain hormones, notably abscisic acid, which inhibits germination, and gibberellin, which ends seed dormancy.
Roots
First to emerge from the seed coat is the primary root, or radicle as it is called in the embryo. This radicle is composed of one major root that is thicker at its base and tapers toward the growing tip. Early root growth is one of the functions of the apical meristem located near the tip of the root. The meristem cells more or less continuously divide, producing more meristem, root cap cells (these are sacrificed to protect the meristem), and undifferentiated root cells. The latter become the primary tissues of the root, first undergoing elongation. Gradually these cells differentiate and mature into specialized cells of root tissues.
{kind=link}
{kind=link}
After the embryonic seedling emerges from the seed coat and starts growing roots and leaves, the embryonic seedling's food reserves are quickly exhausted. At this point photosynthesis provides the energy needed for continuing growth and the seedling now requires a continuous supply of water, nutrients, and light. Most tree species grow a taproot from the seed until moisture or an obstacle is reached and the seedling can then focus on spreading structural roots, absorbing water and nutrients, and letting the leaves manufacture food.
{kind=link}
Tree roots will generally grow in any direction where the correct environment of air, mineral nutrients, and water exists to meet the tree's needs. Roots will stop growing toward dry or poor soil conditions.
Vegetative Reproduction
Besides seeds, trees can be propagated with the assistance of mankind.
* Cultivars are propagated by grafting a bud or stem of the parent tree onto a seedling rootstock of the species.
* Clones are propagated by encouraging roots to develop on cuttings of the parent tree.
Sprouts and suckers start with dormant buds that "come alive" to form new shoots off the parent trees. Sprouts are shoots from stumps of a tree. Suckers are shoots that originate from buds on the root systems. Both sprouts and suckers have the potential to grow into new full size trees genetically identical to the parent. Often times, sprouts and suckers will not grow until the parent tree dies or becomes very sick or suffers from a severe injury. The buds are held in dormancy by hormones produced in the leaves. When these hormone levels drops below a certain point because of increased light level as the old tree thins its branches and leaves, the dormant buds begin to grow.
Vegetative layering is an uncommon form of vegetative propagation. When branches or stems come in contact with the soil, the cambium tissue sometimes will form roots. In this way, former branches might become trunks of several "new" trees genetically identical to the parent.
Tissue culture is an invention of modern science to extract cells of desirable trees and grow these cells into trees within laboratory conditions. Trees from this process are genetically identical to the parent.
Tree Root Physiology
The development of a root system is dependent upon the tree genetics, the soils, and the environment. Root physiology is the ability of the roots to sense the environment and adjust accordingly. Roots develop internally rather than from buds that occur on stems and the parts of the tree above the ground.
Root Parts
The root has several parts summarized below and described in more detail in Topic #1. They are listed from the youngest part of a root to the oldest:
Root Cap – This part of the root is at its very tip. The cap protects the tip of the root as it is forced through the soil by the elongating root tissue behind it. The tip has sensors that can detect rocks and other obstacles in its path growth.
Apical Meristem – This part of the root provides the cells for the root cap in front and for the region of elongation behind this area. The sensors will cause the apical meristem to change directions of growth to avoid an obstacle in its path.
Region of Elongation – Cells in their fixed positions elongate to reach mature dimensions in the region of elongation. A special cell called the vacuole plays a major role in this process. It uses water pressure to push against the walls of cells to stretch the cellulose fibers in the new cells to make them elongate and push the cap through the soil.
Region of Root Hairs – It is in this area where cells develop a more mature form and change into the epidermis and the cortex.
Rhizosphere – The thin layer of soil that sticks to the roots is called the rhizosphere. It is the zone, about one millimeter in width, surrounding the epidermis of living root hairs that contains millions more soil microorganisms than anywhere else in the soil.
Mycorrhizae – Mycorrhizae are beneficial fungi to trees. Mycorrhizal fungi are naturally occurring fungi that prefer the soil in the forest. Mycorrhizae (my-ka-RY-zee) are not exactly a root, and they are not exactly a fungus; they fall halfway between. They are important because they form a symbiotic relationship with a tree's roots and they act as extensions of the roots. They translocate nutrients and moisture back to the root for absorption and use by the tree. In exchange, the tree provides the fungus with carbohydrates, vitamins, and amino acids from photosynthesis. For more information about mycorrhizae, see Topic #1.
The development of a root system is dependent upon the tree genetics, the soils, and the environment. Root physiology is the ability of the roots to sense the environment and adjust accordingly. Roots develop internally rather than from buds that occur on stems and the parts of the tree above the ground.
Root Parts
The root has several parts summarized below and described in more detail in Topic #1. They are listed from the youngest part of a root to the oldest:
Root Cap – This part of the root is at its very tip. The cap protects the tip of the root as it is forced through the soil by the elongating root tissue behind it. The tip has sensors that can detect rocks and other obstacles in its path growth.
Apical Meristem – This part of the root provides the cells for the root cap in front and for the region of elongation behind this area. The sensors will cause the apical meristem to change directions of growth to avoid an obstacle in its path.
Region of Elongation – Cells in their fixed positions elongate to reach mature dimensions in the region of elongation. A special cell called the vacuole plays a major role in this process. It uses water pressure to push against the walls of cells to stretch the cellulose fibers in the new cells to make them elongate and push the cap through the soil.
Region of Root Hairs – It is in this area where cells develop a more mature form and change into the epidermis and the cortex.
Rhizosphere – The thin layer of soil that sticks to the roots is called the rhizosphere. It is the zone, about one millimeter in width, surrounding the epidermis of living root hairs that contains millions more soil microorganisms than anywhere else in the soil.
Mycorrhizae – Mycorrhizae are beneficial fungi to trees. Mycorrhizal fungi are naturally occurring fungi that prefer the soil in the forest. Mycorrhizae (my-ka-RY-zee) are not exactly a root, and they are not exactly a fungus; they fall halfway between. They are important because they form a symbiotic relationship with a tree's roots and they act as extensions of the roots. They translocate nutrients and moisture back to the root for absorption and use by the tree. In exchange, the tree provides the fungus with carbohydrates, vitamins, and amino acids from photosynthesis. For more information about mycorrhizae, see Topic #1.
Tree Trunk Physiology
The trunk supports the crown and holds the leaves up to the light to allow photosynthesis to occur. The trunk is a highly ordered arrangement of living, dying, and dead cells. The roots are bathed in a nutrient rich liquid which transports these nutrients plus moisture to the top of the tree where it is all consumed during photosynthesis. The tree trunk has to expand in diameter as the tree grows in its search for moisture and sunlight.
A tree (and the cells) supports an ever-flowing wet system that must be maintained at all times. If the process fails to provide water at any point, the tree will eventually die due to the failure of both water and food requirements that are necessary for life. For additional information, see the article "The Bark of a Tree".
Outer Bark
The outer bark is a layer of dead cells which protects and covers the layer of living cells of the cambium. Bark is a tree's natural armor and protects it from external threats. Bark also has several physical functions; one is disposing the tree of wastes by absorbing them and locking them into its dead cells and resins in the bark. The bark also influences light passage, it regulates gas exchange, it recognizes and defends against pathogens, it controls tree temperature, and many other things that contribute to the survival of the tree.
Bark textures are relatively uniform according to tree species and the bark makes a great visual marker for tree identification. Bark textures are divided into at least 18 types, that range from smooth (like a beech or Fagus) to spiny (like a locust or Robinia).
Inner Bark or Cork Cambium
This tissue occupies the space between the outer bark and the phloem tissues and is called the cork cambium layer. This is the layer that generates new bark. In roots the cambium tissue may store sugars or starches to fuel the spring sap flow. In leaves, the cambium tissue is the mesophyll cells doing the photosynthesis.
Phloem
Phloem, next to the inner bark, develops from the outside layer of the cambium and is the food track to the roots. Phloem carries large quantities of manufactured food or sugars that are transported from leaves toward roots. When the tree is healthy and growing and sugars are abundant, the extra sugars are stored in the form of starch which can be converted back into sugars and moved by the phloem to where it is needed in the tree.
Cambium
The cambium and its "zone" is a cell generator (reproductive tissue called growth meristem) that produces both the inner bark cells of the phloem and new living wood cells in the xylem. The cambium is a watery layer only a few cells thick.
Xylem
Xylem is living "sapwood" and is located inside the cambial zone and next to the heartwood. Xylem carries water, substances dissolved in water, and minerals from the roots to the leaves. The outer portion of xylem is conducting and storing starch in the symplast plus it conducts water and substances dissolved in water to the leaves. The major structures for water transport in xylem are called vessels in angiosperms (hardwoods) and tracheids in gymnosperms (conifers).
Heartwood
The inner portion of the xylem is non-conducting wood that stores starch and is sometimes called the heartwood. Most of the heartwood is dead woody tissue and serves as a repository for many waste products of the tree's living tissue. It also functions to support the weight of the crown. The cells produced in the spring, called “spring wood”, when the tree is growing quickly, are wide to allow them to carry the large amounts of the water needed for the tree’s rapid growth. Later in the season, the cells produced are smaller, thicker walled and slower growing and called “summer wood”. Spring wood is light in color and fast growing wood, while summer wood is darker and the slower growing wood. This difference in the cells is seen in a cross section of the trunk as bands or rings. From this annual pattern the age of the tree can be counted. These annual rings are typical in temperate forest trees and tropical forest trees that have regular, annual dry seasons. In tropical humid rainforests, trees grow continually and do not have rings. The oldest portion of a tree is at the bottom and at the innermost rings on the trunk.
Symplast
Symplast is the network of living cells and the connections between living cells. Starch is stored in the symplast. Axial parenchyma, ray parenchyma, sieve tubes, companion cells, cork cambium, the cambium, and plasmodesmata all make up the symplast.
Vessels and Tracheids
Vessels (in hardwoods) and tracheids (in conifers) also conduct water and substances dissolved in water. Vessels are vertically aligned tubes made up of dead cells that transport liquid. Vessels are found only in angiosperms. Tracheids are dead, single-celled "pipes" that act much like vessels but are only found in gymnosperms.
Apical Dominance
Apical dominance is the phenomenon whereby the main central stem of the tree is dominant over side branches. The apical bud subjugates all other side branches by producing growth hormones that reach and affect all parts of the tree. Those hormone regulated side branches just grow ever outward and as a result they typically have wide angles of attachment to the stem. Because of this, the apical bud is the only one that grows consistently upward or towards the light while the remaining branches are cast in a supportive role by not being allowed to compete for upward growth.
The trunk supports the crown and holds the leaves up to the light to allow photosynthesis to occur. The trunk is a highly ordered arrangement of living, dying, and dead cells. The roots are bathed in a nutrient rich liquid which transports these nutrients plus moisture to the top of the tree where it is all consumed during photosynthesis. The tree trunk has to expand in diameter as the tree grows in its search for moisture and sunlight.
A tree (and the cells) supports an ever-flowing wet system that must be maintained at all times. If the process fails to provide water at any point, the tree will eventually die due to the failure of both water and food requirements that are necessary for life. For additional information, see the article "The Bark of a Tree".
Outer Bark
The outer bark is a layer of dead cells which protects and covers the layer of living cells of the cambium. Bark is a tree's natural armor and protects it from external threats. Bark also has several physical functions; one is disposing the tree of wastes by absorbing them and locking them into its dead cells and resins in the bark. The bark also influences light passage, it regulates gas exchange, it recognizes and defends against pathogens, it controls tree temperature, and many other things that contribute to the survival of the tree.
Bark textures are relatively uniform according to tree species and the bark makes a great visual marker for tree identification. Bark textures are divided into at least 18 types, that range from smooth (like a beech or Fagus) to spiny (like a locust or Robinia).
Inner Bark or Cork Cambium
This tissue occupies the space between the outer bark and the phloem tissues and is called the cork cambium layer. This is the layer that generates new bark. In roots the cambium tissue may store sugars or starches to fuel the spring sap flow. In leaves, the cambium tissue is the mesophyll cells doing the photosynthesis.
Phloem
Phloem, next to the inner bark, develops from the outside layer of the cambium and is the food track to the roots. Phloem carries large quantities of manufactured food or sugars that are transported from leaves toward roots. When the tree is healthy and growing and sugars are abundant, the extra sugars are stored in the form of starch which can be converted back into sugars and moved by the phloem to where it is needed in the tree.
Cambium
The cambium and its "zone" is a cell generator (reproductive tissue called growth meristem) that produces both the inner bark cells of the phloem and new living wood cells in the xylem. The cambium is a watery layer only a few cells thick.
Xylem
Xylem is living "sapwood" and is located inside the cambial zone and next to the heartwood. Xylem carries water, substances dissolved in water, and minerals from the roots to the leaves. The outer portion of xylem is conducting and storing starch in the symplast plus it conducts water and substances dissolved in water to the leaves. The major structures for water transport in xylem are called vessels in angiosperms (hardwoods) and tracheids in gymnosperms (conifers).
Heartwood
The inner portion of the xylem is non-conducting wood that stores starch and is sometimes called the heartwood. Most of the heartwood is dead woody tissue and serves as a repository for many waste products of the tree's living tissue. It also functions to support the weight of the crown. The cells produced in the spring, called “spring wood”, when the tree is growing quickly, are wide to allow them to carry the large amounts of the water needed for the tree’s rapid growth. Later in the season, the cells produced are smaller, thicker walled and slower growing and called “summer wood”. Spring wood is light in color and fast growing wood, while summer wood is darker and the slower growing wood. This difference in the cells is seen in a cross section of the trunk as bands or rings. From this annual pattern the age of the tree can be counted. These annual rings are typical in temperate forest trees and tropical forest trees that have regular, annual dry seasons. In tropical humid rainforests, trees grow continually and do not have rings. The oldest portion of a tree is at the bottom and at the innermost rings on the trunk.
Symplast
Symplast is the network of living cells and the connections between living cells. Starch is stored in the symplast. Axial parenchyma, ray parenchyma, sieve tubes, companion cells, cork cambium, the cambium, and plasmodesmata all make up the symplast.
Vessels and Tracheids
Vessels (in hardwoods) and tracheids (in conifers) also conduct water and substances dissolved in water. Vessels are vertically aligned tubes made up of dead cells that transport liquid. Vessels are found only in angiosperms. Tracheids are dead, single-celled "pipes" that act much like vessels but are only found in gymnosperms.
Apical Dominance
Apical dominance is the phenomenon whereby the main central stem of the tree is dominant over side branches. The apical bud subjugates all other side branches by producing growth hormones that reach and affect all parts of the tree. Those hormone regulated side branches just grow ever outward and as a result they typically have wide angles of attachment to the stem. Because of this, the apical bud is the only one that grows consistently upward or towards the light while the remaining branches are cast in a supportive role by not being allowed to compete for upward growth.
Physiology of Leaves
A tree crown is where most bud formation takes place. The tree bud is simply a small bundle of growing tissue which develops into embryonic leaves, flowers and shoots and is essential for primary tree crown and canopy growth. In addition to branch growth, buds are responsible for flower formation and leaf production. A tree's small budding structure is wrapped in a simple protecting leaf called the cataphylls. These protected buds allow all plants to continue to grow and produce tiny new leaves and flowers even when environmental conditions are adverse or limiting.
Like roots and trunks, branches grow in length from growth cells that make up the meristematic tissues which are contained in growing buds. This limb and branch bud growth determine a tree’s crown shape, size and height. The tree crown's central and terminal leader grows from a bud cell called the apical meristem which determines the tree’s height.
Some buds contain tiny preformed flowers, or both leaves and flowers. Buds may be terminal (on the end of the shoot) or lateral (on the side of the shoot, usually at the base of the leaves).
Chlorophyll
Chlorophyll is the chemical compound where solar energy (light) is captured and photosynthesis happens. Chlorophyll is continuously produced and broken down during the growing season. The heart of the chlorophyll compound is a magnesium molecule. The magnesium molecule is bonded to many molecules of hydrogen, carbon, oxygen, and nitrogen.
Photosynthesis
All trees and most plants photosynthesize and respire. Photosynthesis is a process unique to green plants and produces sugars, which are "tree food". Trees produce their own food called sugar. These sugars are not like the refined sugar that people eat, and these sugars don’t usually taste sweet, but the basic organic components are similar. Unlike trees, animals only respire and cannot produce their own food.
The chemical formula for producing sugars is:
(6) CO2 + (6) H2O + sunlight energy produces C6H12O6 + (6) O2
or
Carbon dioxide plus water plus sunlight energy produces glucose and oxygen
Photosynthesis literally means “to put together with light”. It occurs only in the chloroplasts which are tiny sub-cellular structures contained in the cells of leaves and green stems. Inside the chloroplast, the chlorophyll molecules use energy from the sun to make the hydrogen from the water combined with the carbon and oxygen and from the soluble carbohydrate to produce sugars. The sugars can then be made into starch for storage in the wood.
Energy is stored in the bonds of sugar molecules such as glucose and fructose. Oxygen is a by-product of photosynthesis. The oxygen molecules produced by photosynthesis are not necessarily the same oxygen molecules the plants use for respiration.
Respiration
Trees need energy to grow. This energy is released from the food made by photosynthesis in a process called respiration. In respiration, trees convert the sugars (photosynthates) back into energy for growth and other life and metabolic processes. The released energy drives a variety of metabolic actions. It is the same process that animals use when they respire. So, either the plant uses its own stored sugars, or some animal (or decomposer) consumes the plant, and uses the stored sugars. In either case, the sugars are valued chemicals because they contain energy, as well as important elements (carbon, hydrogen, and oxygen).
Chemically speaking, the process is similar to the oxidation that occurs as wood is burned, producing heat. When compounds combine with oxygen, the process is often referred to as “burning”, for example, athlete’s “burn” energy (sugars) as they exercise. The harder they exercise, the more sugars they burn so the more oxygen they need. That is why at full speed, they are breathing very fast. Athletes take up oxygen through their lungs. Trees take up oxygen in the air through the stomata in their leaves and oxygen in the soil pores through their roots.
A tree will draw nutrients and minerals from the soil, break them down and put them back together to form compounds and chemicals that become a growing tree. The most common material made by a tree is cellulose. Cellulose is a complex sugar that is the main component of wood and many other plant tissues. It is also an extremely useful material for human uses, such as food products, paper, strengthener in plastics and concrete, clothing, and other things.
Transpiration Stomata
Transpiration is the process of water movement through a tree. Water enters the tree through its root hairs. The process of osmosis moves the water from the root hair to the root cortex cells and then to the xylem vessels where it is transported to the leaves for use in cooling the tree primarily and also for the process of photosynthesis. From this point, the water is sent back to the roots or released from the leaf through the stomata by the guard cells where it can actually cool the tree in the spongy meseophyll cells and the tree as the water evaporates.
Guard cells are cells surrounding each stomata (which is a type of hole in the leaf surface). They help to regulate the rate of transpiration by opening and closing the stomata. When the guard cells are turgid, or swollen, and the stomatal opening is large, the water potential of the guard cells drops and water enters the guard cells. This turgidity is caused by the accumulation of K+ (potassium ions) in the guard cells. When the guard cells have lost water, the cells become flaccid and the stomatal opening closes. This may occur when the plant has lost an excessive amount of water. In addition, it generally occurs daily as light levels drop and the use of carbon dioxide in photosynthesis decreases.
As water vapor evaporates from the stomata it is replaced by water being pulled from the xylem. The water in the xylem is coming from the tree’s roots. In other words, water is being pulled up through the tree by transpiration, like drinking water through a straw. Transpiration uses about 90% of the water that enters the tree. The other 10% is an ingredient in photosynthesis and cell growth and it is called the tree’s sap.
Transpiration serves three essential roles:
1. Movement of minerals – water moves up from the root, by way of the xylem, and sugars which are
products of photosynthesis throughout the tree by way of the phloem. Water serves as both the solvent
and the avenue of transport.
2. Cooling – 80% of the cooling effect of a shade tree comes from the evaporative cooling effects of
transpiration. This benefits both plants and humans.
3. Turgor pressure – Water maintains the turgor pressure in cells much like air inflates a balloon, giving the
non-woody plant parts form. Turgidity is important so the plant can remain stiff and upright and gain a
competitive advantage when it comes to sunlight. Turgidity is also important for the functioning of the guard
cells, which surround the stomata and regulate water loss and carbon dioxide uptake. Turgidity also is the
force that pushes roots through the soil.
A tree crown is where most bud formation takes place. The tree bud is simply a small bundle of growing tissue which develops into embryonic leaves, flowers and shoots and is essential for primary tree crown and canopy growth. In addition to branch growth, buds are responsible for flower formation and leaf production. A tree's small budding structure is wrapped in a simple protecting leaf called the cataphylls. These protected buds allow all plants to continue to grow and produce tiny new leaves and flowers even when environmental conditions are adverse or limiting.
Like roots and trunks, branches grow in length from growth cells that make up the meristematic tissues which are contained in growing buds. This limb and branch bud growth determine a tree’s crown shape, size and height. The tree crown's central and terminal leader grows from a bud cell called the apical meristem which determines the tree’s height.
Some buds contain tiny preformed flowers, or both leaves and flowers. Buds may be terminal (on the end of the shoot) or lateral (on the side of the shoot, usually at the base of the leaves).
Chlorophyll
Chlorophyll is the chemical compound where solar energy (light) is captured and photosynthesis happens. Chlorophyll is continuously produced and broken down during the growing season. The heart of the chlorophyll compound is a magnesium molecule. The magnesium molecule is bonded to many molecules of hydrogen, carbon, oxygen, and nitrogen.
Photosynthesis
All trees and most plants photosynthesize and respire. Photosynthesis is a process unique to green plants and produces sugars, which are "tree food". Trees produce their own food called sugar. These sugars are not like the refined sugar that people eat, and these sugars don’t usually taste sweet, but the basic organic components are similar. Unlike trees, animals only respire and cannot produce their own food.
The chemical formula for producing sugars is:
(6) CO2 + (6) H2O + sunlight energy produces C6H12O6 + (6) O2
or
Carbon dioxide plus water plus sunlight energy produces glucose and oxygen
Photosynthesis literally means “to put together with light”. It occurs only in the chloroplasts which are tiny sub-cellular structures contained in the cells of leaves and green stems. Inside the chloroplast, the chlorophyll molecules use energy from the sun to make the hydrogen from the water combined with the carbon and oxygen and from the soluble carbohydrate to produce sugars. The sugars can then be made into starch for storage in the wood.
Energy is stored in the bonds of sugar molecules such as glucose and fructose. Oxygen is a by-product of photosynthesis. The oxygen molecules produced by photosynthesis are not necessarily the same oxygen molecules the plants use for respiration.
Respiration
Trees need energy to grow. This energy is released from the food made by photosynthesis in a process called respiration. In respiration, trees convert the sugars (photosynthates) back into energy for growth and other life and metabolic processes. The released energy drives a variety of metabolic actions. It is the same process that animals use when they respire. So, either the plant uses its own stored sugars, or some animal (or decomposer) consumes the plant, and uses the stored sugars. In either case, the sugars are valued chemicals because they contain energy, as well as important elements (carbon, hydrogen, and oxygen).
Chemically speaking, the process is similar to the oxidation that occurs as wood is burned, producing heat. When compounds combine with oxygen, the process is often referred to as “burning”, for example, athlete’s “burn” energy (sugars) as they exercise. The harder they exercise, the more sugars they burn so the more oxygen they need. That is why at full speed, they are breathing very fast. Athletes take up oxygen through their lungs. Trees take up oxygen in the air through the stomata in their leaves and oxygen in the soil pores through their roots.
A tree will draw nutrients and minerals from the soil, break them down and put them back together to form compounds and chemicals that become a growing tree. The most common material made by a tree is cellulose. Cellulose is a complex sugar that is the main component of wood and many other plant tissues. It is also an extremely useful material for human uses, such as food products, paper, strengthener in plastics and concrete, clothing, and other things.
Transpiration Stomata
Transpiration is the process of water movement through a tree. Water enters the tree through its root hairs. The process of osmosis moves the water from the root hair to the root cortex cells and then to the xylem vessels where it is transported to the leaves for use in cooling the tree primarily and also for the process of photosynthesis. From this point, the water is sent back to the roots or released from the leaf through the stomata by the guard cells where it can actually cool the tree in the spongy meseophyll cells and the tree as the water evaporates.
Guard cells are cells surrounding each stomata (which is a type of hole in the leaf surface). They help to regulate the rate of transpiration by opening and closing the stomata. When the guard cells are turgid, or swollen, and the stomatal opening is large, the water potential of the guard cells drops and water enters the guard cells. This turgidity is caused by the accumulation of K+ (potassium ions) in the guard cells. When the guard cells have lost water, the cells become flaccid and the stomatal opening closes. This may occur when the plant has lost an excessive amount of water. In addition, it generally occurs daily as light levels drop and the use of carbon dioxide in photosynthesis decreases.
As water vapor evaporates from the stomata it is replaced by water being pulled from the xylem. The water in the xylem is coming from the tree’s roots. In other words, water is being pulled up through the tree by transpiration, like drinking water through a straw. Transpiration uses about 90% of the water that enters the tree. The other 10% is an ingredient in photosynthesis and cell growth and it is called the tree’s sap.
Transpiration serves three essential roles:
1. Movement of minerals – water moves up from the root, by way of the xylem, and sugars which are
products of photosynthesis throughout the tree by way of the phloem. Water serves as both the solvent
and the avenue of transport.
2. Cooling – 80% of the cooling effect of a shade tree comes from the evaporative cooling effects of
transpiration. This benefits both plants and humans.
3. Turgor pressure – Water maintains the turgor pressure in cells much like air inflates a balloon, giving the
non-woody plant parts form. Turgidity is important so the plant can remain stiff and upright and gain a
competitive advantage when it comes to sunlight. Turgidity is also important for the functioning of the guard
cells, which surround the stomata and regulate water loss and carbon dioxide uptake. Turgidity also is the
force that pushes roots through the soil.
Water movement in trees is also a factor of osmotic pressure and capillary action. Osmotic pressure is defined as water flowing through a permeable membrane in the direction of a higher salt concentration. Water will continue to flow in the direction of the highest salt concentration until the salts have been diluted to the point that the concentrations on both sides of the membrane are equal.
Capillary action refers to the chemical forces that move water as a continuous film rather than as individual molecules. Water molecules in the soil and in the tree cling to one another. So when one molecule is drawn up the plant stem, it pulls another one along with it. These forces that link water molecules together can be overcome by gravity.
Glucose
After photosynthesis produces the sugar glucose, the energy in it is used by trees to drive metabolic processes that produce tissues and maintain life functions.
Distribution of Glucose
A tree will typically move energy according to these priorities, listed in order. As energy in the form of glucose becomes limited, a tree will begin to reduce resources beginning with the lowest item on this list and working its way up.
Capillary action refers to the chemical forces that move water as a continuous film rather than as individual molecules. Water molecules in the soil and in the tree cling to one another. So when one molecule is drawn up the plant stem, it pulls another one along with it. These forces that link water molecules together can be overcome by gravity.
Glucose
After photosynthesis produces the sugar glucose, the energy in it is used by trees to drive metabolic processes that produce tissues and maintain life functions.
Distribution of Glucose
A tree will typically move energy according to these priorities, listed in order. As energy in the form of glucose becomes limited, a tree will begin to reduce resources beginning with the lowest item on this list and working its way up.
- Maintain respiration of all parts.
- Produce fine roots and leaves.
- Produce flowers and seed.
- Extend branches and roots.
- Store energy rich chemicals.
- Add wood to stem, roots, and branches.
- Create anti-pest chemicals for defense.
A tree with a diminishing crown will become more vulnerable to insects and diseases rather quickly. That’s one reason why arborists are so keen to maintain a vigorous growing environment.
Branches
Branch Growth
A tree branch is a woody structural member connected to but not part of the central trunk of a tree. Its purpose is to hold the leaves up to the sunlight. While branches can be nearly horizontal, vertical, or diagonal, the majority of trees have upwardly diagonal branches. Large branches are known as boughs and small branches are known as twigs. The term "bough" refers only to branches coming directly from the trunk, while "twig" often refers to a terminus. A crotch or fork is an area where a trunk splits into two or more boughs. Branches grow in annual increments. Those increments begin at nodes from which a twig will grow and eventually become a bough in subsequent seasons.
Knots in sawn timber are the result of branches. The trunk, with its large diameter, is the source of timber logs. To get clear wood or timber without knots, the branches should be pruned off at an early age so as the wood grows over the branch stumps the rest of the tree’s wood is clear of knots.
Evolutionary Design
Trees are relatively stable structures in a world where climatic factors and fluctuations test them all the time. In response to these fluctuations trees do not grow ever stronger to overcome the structural stresses that they periodically endure. Instead, trees have been designed by evolution to react passively to extreme climatic events and progressive wind pruning. The tree fails by allowing the branches at the weakest crotches to break off from the outer edge of the tree crown. These crotches have variable strengths specifically for facilitating subtle and progressive branch failure. The successive years of growth create more branch unions of variable strength. As they do that, the remaining structural elements become stronger and the likelihood of the whole tree blowing over is reduced. These variable strengths appear to facilitate wind pruning and give the tree the ability to adapt and change its size and shape rather than simply failing at the roots. In other words, trees reduce structural stress in a controlled way by creating some relatively weak branch unions throughout the canopy of mature trees. These potential pruning points enable the tree to survive a storm rather than becoming a total failure.
While arborists and landscape architects think these relatively weak and "acute" crotches are a problem, we now realize that for the tree, they are a benefit because they are all a tree needs to passively survive a strong wind. Arborists should use this information when trimming trees to be sure they are leaving small wounds that grow over quickly rather than natural rip wounds that can be extensive and are therefore more likely to lead to decay. The crucial thing is that by closely mimicking how a tree is designed by evolution to be wind pruned means that this type of pruning on landscape trees can reduce risks while also keeping the tree looking as natural as possible.
Co-dominant Stems
Co-dominant stems are created when a branch sends out two identical branches. They both have their own apical buds, both leaders have the same resources, and they both grow at the same rate. Because they set off at the same point in time and they are genetically identical, one cannot outgrow the other. The result of such co-dominant morphology is inevitably strong competition between the two apically dominant buds, and that means strong growth from a weak crotch.
Because the two leaders rise closely to each other, the bark becomes included in the acutely angled crotch and this prevents the wood fibers of the two leaders from binding properly with each other. The final problem comes when high winds act on the crotch and the leaders move back and forth from each other. This can cause a crack to form at the base of the acute crotch. The crack expands down the branch further weakening the crotch and eventually resulting in one or both of the leaders breaking and falling off the tree.
Acute Unions
Wider than the co-dominant stems are acute unions. Acute crotches have been extensively studied and as a result are widely understood to be stronger than the co-dominant crotch and therefore are not likely to be a point at which the branch will fail in high winds. As a result, acute crotches are also considered to be desirable. The degree of relative strength goes up with the relative widening of the angle between the branches. This is what the arborist should look for when pruning to leave a tree with minimal wounds and a natural appearance.
Sway Factor
By pruning a number of these primary branches with weak crotches, back to one of the stronger branch unions, not only is the tree’s shape reduced, but the tree's peripheral mass is also reduced slightly. This reduced mass, reduces the amount of time it takes for the tree to sway and recover after being hit by a gust of wind. The removal of some peripheral mass will raise the sway frequency by a significant percentage greater than the simple percentage of foliage removed. This reduced mass also means that the tree is less likely to fail in total.
Emergence Above the Forest Canopy
When a tree falls in the forest, the extra light on the forest floor encourages seeds to germinate and the seedlings grow rapidly to take advantage of this light. The tree grows very rapidly to fill the space vacated by a tree failure. As the tree grows as tall as the surrounding forest trees, it emerges into the area above the general height of the surrounding forest trees it becomes exposed to wind or frosts and a full range of adverse occurrences. The tree has little choice because the growth hormones produced by the apical bud to continue the upward mode of growth. It is hormonally compelled to grow ever upward.
If the growth tip and the apical bud become damaged or knocked off, it triggers some profound changes in the structure of the tree. For the first time and in just one brief moment in its life, there is no apical bud and therefore no hormones to impose apical dominance on all of the other lateral branches. The side branches have a very brief opportunity to grow in forms that are less rigorously controlled than was the case in the young forest tree. So in addition to the tip forking, many of the lateral branches also form in this moment and a more rounded shape develops at the top of the tree.
Crown Formation
Most trees have a crown that is typical of the species. For example, most crabapples have a round form, making the tree look like a lollypop. However there are some crabapples that have been selected for its genetic ability to develop a columnar form or a vase shape or a weeping form.
Aside from this genetic ability to generate a tree crown form, some trees have unexpected forms that occur during the tree’s development. These changes need a trigger that activates some or all of the numerous apical buds that replace the single apical bud that became damaged. The trigger can be a late spring frost or a bird that lands on the bud and it breaks off just as the succulent leaves are emerging. This will cause the peripheral buds to become active and result in weak crotches and a random form not typical of the species or cultivar.
Mid-summer droughts can also stress the tree and cause it to die back from the tips of the branches so as to make the leaf area smaller which enables the tree to survive the drought and grow on to maturity. Other natural causes include pests, root damage or loss, and certain diseases.
Branch Growth
A tree branch is a woody structural member connected to but not part of the central trunk of a tree. Its purpose is to hold the leaves up to the sunlight. While branches can be nearly horizontal, vertical, or diagonal, the majority of trees have upwardly diagonal branches. Large branches are known as boughs and small branches are known as twigs. The term "bough" refers only to branches coming directly from the trunk, while "twig" often refers to a terminus. A crotch or fork is an area where a trunk splits into two or more boughs. Branches grow in annual increments. Those increments begin at nodes from which a twig will grow and eventually become a bough in subsequent seasons.
Knots in sawn timber are the result of branches. The trunk, with its large diameter, is the source of timber logs. To get clear wood or timber without knots, the branches should be pruned off at an early age so as the wood grows over the branch stumps the rest of the tree’s wood is clear of knots.
Evolutionary Design
Trees are relatively stable structures in a world where climatic factors and fluctuations test them all the time. In response to these fluctuations trees do not grow ever stronger to overcome the structural stresses that they periodically endure. Instead, trees have been designed by evolution to react passively to extreme climatic events and progressive wind pruning. The tree fails by allowing the branches at the weakest crotches to break off from the outer edge of the tree crown. These crotches have variable strengths specifically for facilitating subtle and progressive branch failure. The successive years of growth create more branch unions of variable strength. As they do that, the remaining structural elements become stronger and the likelihood of the whole tree blowing over is reduced. These variable strengths appear to facilitate wind pruning and give the tree the ability to adapt and change its size and shape rather than simply failing at the roots. In other words, trees reduce structural stress in a controlled way by creating some relatively weak branch unions throughout the canopy of mature trees. These potential pruning points enable the tree to survive a storm rather than becoming a total failure.
While arborists and landscape architects think these relatively weak and "acute" crotches are a problem, we now realize that for the tree, they are a benefit because they are all a tree needs to passively survive a strong wind. Arborists should use this information when trimming trees to be sure they are leaving small wounds that grow over quickly rather than natural rip wounds that can be extensive and are therefore more likely to lead to decay. The crucial thing is that by closely mimicking how a tree is designed by evolution to be wind pruned means that this type of pruning on landscape trees can reduce risks while also keeping the tree looking as natural as possible.
Co-dominant Stems
Co-dominant stems are created when a branch sends out two identical branches. They both have their own apical buds, both leaders have the same resources, and they both grow at the same rate. Because they set off at the same point in time and they are genetically identical, one cannot outgrow the other. The result of such co-dominant morphology is inevitably strong competition between the two apically dominant buds, and that means strong growth from a weak crotch.
{kind=link}
Because the two leaders rise closely to each other, the bark becomes included in the acutely angled crotch and this prevents the wood fibers of the two leaders from binding properly with each other. The final problem comes when high winds act on the crotch and the leaders move back and forth from each other. This can cause a crack to form at the base of the acute crotch. The crack expands down the branch further weakening the crotch and eventually resulting in one or both of the leaders breaking and falling off the tree.
Acute Unions
Wider than the co-dominant stems are acute unions. Acute crotches have been extensively studied and as a result are widely understood to be stronger than the co-dominant crotch and therefore are not likely to be a point at which the branch will fail in high winds. As a result, acute crotches are also considered to be desirable. The degree of relative strength goes up with the relative widening of the angle between the branches. This is what the arborist should look for when pruning to leave a tree with minimal wounds and a natural appearance.
Sway Factor
By pruning a number of these primary branches with weak crotches, back to one of the stronger branch unions, not only is the tree’s shape reduced, but the tree's peripheral mass is also reduced slightly. This reduced mass, reduces the amount of time it takes for the tree to sway and recover after being hit by a gust of wind. The removal of some peripheral mass will raise the sway frequency by a significant percentage greater than the simple percentage of foliage removed. This reduced mass also means that the tree is less likely to fail in total.
Emergence Above the Forest Canopy
When a tree falls in the forest, the extra light on the forest floor encourages seeds to germinate and the seedlings grow rapidly to take advantage of this light. The tree grows very rapidly to fill the space vacated by a tree failure. As the tree grows as tall as the surrounding forest trees, it emerges into the area above the general height of the surrounding forest trees it becomes exposed to wind or frosts and a full range of adverse occurrences. The tree has little choice because the growth hormones produced by the apical bud to continue the upward mode of growth. It is hormonally compelled to grow ever upward.
If the growth tip and the apical bud become damaged or knocked off, it triggers some profound changes in the structure of the tree. For the first time and in just one brief moment in its life, there is no apical bud and therefore no hormones to impose apical dominance on all of the other lateral branches. The side branches have a very brief opportunity to grow in forms that are less rigorously controlled than was the case in the young forest tree. So in addition to the tip forking, many of the lateral branches also form in this moment and a more rounded shape develops at the top of the tree.
Crown Formation
Most trees have a crown that is typical of the species. For example, most crabapples have a round form, making the tree look like a lollypop. However there are some crabapples that have been selected for its genetic ability to develop a columnar form or a vase shape or a weeping form.
Aside from this genetic ability to generate a tree crown form, some trees have unexpected forms that occur during the tree’s development. These changes need a trigger that activates some or all of the numerous apical buds that replace the single apical bud that became damaged. The trigger can be a late spring frost or a bird that lands on the bud and it breaks off just as the succulent leaves are emerging. This will cause the peripheral buds to become active and result in weak crotches and a random form not typical of the species or cultivar.
Mid-summer droughts can also stress the tree and cause it to die back from the tips of the branches so as to make the leaf area smaller which enables the tree to survive the drought and grow on to maturity. Other natural causes include pests, root damage or loss, and certain diseases.
Climate's Role in Tree Physiology
Large, healthy, fast-growing trees provide climate benefits faster than small, slow-growing trees. It has to be stressed that because trees might be good in the struggle against climate change, they need to be in good health to maximize gas exchange and produce wood biomass. For city trees, it is important to select a tree to match the city environment in terms of local climate, physiology and growth, in order to obtain the maximum growth and benefits in harsh conditions.
It should be emphasized that a tree growing in the city and battling environmental challenges like those listed below, cannot maximize gas exchange and produce wood biomass. Therefore, tree selection is of utmost importance in assuring that these benefits fully materialize.
Air Temperature
Trees and shrubs obtain optimum growing conditions across a wide range of temperatures but hot temperatures have a negative effect on their physiology. However, trees can dissipate tremendous heat loads if allowed to function normally. Also, higher temperatures may indirectly affect the range and impact of insect species. It is highly probable that insects will be a major problem for city trees under future global warming, and it seems certain that insect distributions will also change.
Growing Season
Shifts in timing and length of the growing season can alter the physiology of trees and interact with their dormancy mechanisms. If the higher temperatures initiate the trees to decrease the tolerance to low temperatures, the trees may be more susceptible to damages from spells of late spring and early autumn frost.
Oxygen Deficiency
Soil oxygen concentration is lower in the soil than the oxygen level in the atmosphere and generally ranges around 16%. When the oxygen content falls below 10%, water uptake by trees is diminished. This is because of reduced tissue permeability and the interruption of root growth. As a consequence, mineral nutrient uptake and the capacity of roots to synthesize growth regulators and nitrogen compounds are also reduced.
Flooding leads to saturated soils but the oxygen deficiency occurs almost immediately. Soil flooding also causes a rapid decrease in photosynthesis and a decrease in the growth rate of the tree. Contrary to this, the effect from compacted soil occurs much more gradually. Furthermore, the aerobic microorganisms are readily replaced by anaerobic ones.
When root growth encounters soil compaction, they produce numerous fine lateral roots which are capable of penetrating the smaller pores and thereby allow for increase in root diameter. However, this action does greatly reduce the growth rate of the tree, so compacted soil should be avoided.
Soil Temperatures
The threshold over which essential physiological processes are slowed down or completely inhibited is 95°F (35°C), especially if other stresses like drought are added to this. To lessen the impact, city trees should be heavily mulched to minimize the heating effect of the paved surface and to allow air and water to reach the roots. Only heat tolerant species and cultivars should be selected for installation in heavily paved areas.
Water Stress
It is known that water stress is the major cause of shortened lifespan and tree mortality in the city. Moisture stress interferes with tree defense systems and increases the likelihood of insect attack. Water stress is extremely dangerous in the first 2 to 3 years after planting. If the quality of plant material is poor, this can result in an unbalanced canopy/root ratio. As a consequence, the transpiration demand exceeds the water up-taking capacity of the roots.
The impacts of climate change on the water supply to the landscape will be significant, but can be reduced by sound water management. Shortages of water, in the summer, can be helped by using stored water and concentrating this water on the most important trees. Irrigation between 9pm and 9am using a control station to schedule irrigation will reduce evaporation losses and provide efficient use of irrigation water. In the long-term, it will become advisable to adopt planting schemes to the new climatic regime of a particular area. For example, select trees tolerant of warmer temperatures when installing new trees. Another example is to reduce evaporation in reservoirs by up to 50% if the shoreline is shaded with trees.
Implication of Climate Change
One important implication of climate change is that the increasing winter temperature will increase the activity and therefore the oxygen demand of trees in winter, so roots may become less tolerant to waterlogging. In general, building a good soil structure will improve the tree root system, especially rooting depth, and make the tree more tolerant to drought.
Nutrients
Trophic conditions of urban soils are extremely variable and nutrient deficiency can sometimes occur in city trees. Phosphorus is generally present but often not available due to insolubility. Potassium is usually present in a relatively good concentration and deficiency symptoms are quite rare. Nitrogen is the element which more frequently limits tree growth in the city. Even if organic matter is present, nitrogen can be deficient when the carbon/nitrogen ratio is high, since nitrogen is needed for the organic matter decomposition by soil microorganisms.
Current tree care recommendations suggest limiting fertilization during the establishment period. Instead, an application of mycorrhizae on the tree’s roots at planting time is much more beneficial.
Other experts believe that growth in the first year can be affected by internal reserves of nutrients and photosynthates as a result of being well fertilized in the nursery. On the other hand the application of inorganic fertilizers aids tree establishment and is an acceptable method of improving tree health and vigor. For example, nitrogen fertilization at the planting time, has positively affected growth in the first year after installation.
Nutrient Absorption
Nutrient uptake by roots in the soil is achieved by cation exchange. Fibrous roots pump positively charged hydrogen ions (H+) into the soil which displace cations attached to negatively charged soil particles, making the cations attached to nutrients, available for uptake by the root. Fibrous roots then become important for the uptake of nutrients as well as water particles.
Solar Radiation
Sunscald occurs, generally, during cold bright days in midwinter. Direct sunrays heat south-facing branches and trunks. The sun raises bark temperature above freezing and causes some cells to become active. When the sun is gone, the active cells freeze and die. This injury does not easily heal and this will result in an interruption of sap flow, which in turn, causes dieback and enhances the sensitivity to parasites, especially in young trees and those recently transplanted. In addition, due to climate change, daytime temperatures are becoming higher and sunnier, meaning that temperatures can more easily reach the threshold after which sunscald becomes more likely.
Root Tolerance
Below ground, trees growing in the city must often tolerate poor, infertile soils (often made up of rubble); compaction, limited soil volumes, drought and poisonous effluents due to de-icing salts, other chemicals and oils, and they are in constant danger from construction events. It is important to remember that trees are long-lived beings, living from a few decades to many hundred years, depending on species. During their lifetime trees will experience great changes in their environment, both climatic and social, and these changes occur continuously.
Large, healthy, fast-growing trees provide climate benefits faster than small, slow-growing trees. It has to be stressed that because trees might be good in the struggle against climate change, they need to be in good health to maximize gas exchange and produce wood biomass. For city trees, it is important to select a tree to match the city environment in terms of local climate, physiology and growth, in order to obtain the maximum growth and benefits in harsh conditions.
It should be emphasized that a tree growing in the city and battling environmental challenges like those listed below, cannot maximize gas exchange and produce wood biomass. Therefore, tree selection is of utmost importance in assuring that these benefits fully materialize.
Air Temperature
Trees and shrubs obtain optimum growing conditions across a wide range of temperatures but hot temperatures have a negative effect on their physiology. However, trees can dissipate tremendous heat loads if allowed to function normally. Also, higher temperatures may indirectly affect the range and impact of insect species. It is highly probable that insects will be a major problem for city trees under future global warming, and it seems certain that insect distributions will also change.
Growing Season
Shifts in timing and length of the growing season can alter the physiology of trees and interact with their dormancy mechanisms. If the higher temperatures initiate the trees to decrease the tolerance to low temperatures, the trees may be more susceptible to damages from spells of late spring and early autumn frost.
Oxygen Deficiency
Soil oxygen concentration is lower in the soil than the oxygen level in the atmosphere and generally ranges around 16%. When the oxygen content falls below 10%, water uptake by trees is diminished. This is because of reduced tissue permeability and the interruption of root growth. As a consequence, mineral nutrient uptake and the capacity of roots to synthesize growth regulators and nitrogen compounds are also reduced.
Flooding leads to saturated soils but the oxygen deficiency occurs almost immediately. Soil flooding also causes a rapid decrease in photosynthesis and a decrease in the growth rate of the tree. Contrary to this, the effect from compacted soil occurs much more gradually. Furthermore, the aerobic microorganisms are readily replaced by anaerobic ones.
When root growth encounters soil compaction, they produce numerous fine lateral roots which are capable of penetrating the smaller pores and thereby allow for increase in root diameter. However, this action does greatly reduce the growth rate of the tree, so compacted soil should be avoided.
Soil Temperatures
The threshold over which essential physiological processes are slowed down or completely inhibited is 95°F (35°C), especially if other stresses like drought are added to this. To lessen the impact, city trees should be heavily mulched to minimize the heating effect of the paved surface and to allow air and water to reach the roots. Only heat tolerant species and cultivars should be selected for installation in heavily paved areas.
Water Stress
It is known that water stress is the major cause of shortened lifespan and tree mortality in the city. Moisture stress interferes with tree defense systems and increases the likelihood of insect attack. Water stress is extremely dangerous in the first 2 to 3 years after planting. If the quality of plant material is poor, this can result in an unbalanced canopy/root ratio. As a consequence, the transpiration demand exceeds the water up-taking capacity of the roots.
The impacts of climate change on the water supply to the landscape will be significant, but can be reduced by sound water management. Shortages of water, in the summer, can be helped by using stored water and concentrating this water on the most important trees. Irrigation between 9pm and 9am using a control station to schedule irrigation will reduce evaporation losses and provide efficient use of irrigation water. In the long-term, it will become advisable to adopt planting schemes to the new climatic regime of a particular area. For example, select trees tolerant of warmer temperatures when installing new trees. Another example is to reduce evaporation in reservoirs by up to 50% if the shoreline is shaded with trees.
Implication of Climate Change
One important implication of climate change is that the increasing winter temperature will increase the activity and therefore the oxygen demand of trees in winter, so roots may become less tolerant to waterlogging. In general, building a good soil structure will improve the tree root system, especially rooting depth, and make the tree more tolerant to drought.
Nutrients
Trophic conditions of urban soils are extremely variable and nutrient deficiency can sometimes occur in city trees. Phosphorus is generally present but often not available due to insolubility. Potassium is usually present in a relatively good concentration and deficiency symptoms are quite rare. Nitrogen is the element which more frequently limits tree growth in the city. Even if organic matter is present, nitrogen can be deficient when the carbon/nitrogen ratio is high, since nitrogen is needed for the organic matter decomposition by soil microorganisms.
Current tree care recommendations suggest limiting fertilization during the establishment period. Instead, an application of mycorrhizae on the tree’s roots at planting time is much more beneficial.
Other experts believe that growth in the first year can be affected by internal reserves of nutrients and photosynthates as a result of being well fertilized in the nursery. On the other hand the application of inorganic fertilizers aids tree establishment and is an acceptable method of improving tree health and vigor. For example, nitrogen fertilization at the planting time, has positively affected growth in the first year after installation.
Nutrient Absorption
Nutrient uptake by roots in the soil is achieved by cation exchange. Fibrous roots pump positively charged hydrogen ions (H+) into the soil which displace cations attached to negatively charged soil particles, making the cations attached to nutrients, available for uptake by the root. Fibrous roots then become important for the uptake of nutrients as well as water particles.
Solar Radiation
Sunscald occurs, generally, during cold bright days in midwinter. Direct sunrays heat south-facing branches and trunks. The sun raises bark temperature above freezing and causes some cells to become active. When the sun is gone, the active cells freeze and die. This injury does not easily heal and this will result in an interruption of sap flow, which in turn, causes dieback and enhances the sensitivity to parasites, especially in young trees and those recently transplanted. In addition, due to climate change, daytime temperatures are becoming higher and sunnier, meaning that temperatures can more easily reach the threshold after which sunscald becomes more likely.
Root Tolerance
Below ground, trees growing in the city must often tolerate poor, infertile soils (often made up of rubble); compaction, limited soil volumes, drought and poisonous effluents due to de-icing salts, other chemicals and oils, and they are in constant danger from construction events. It is important to remember that trees are long-lived beings, living from a few decades to many hundred years, depending on species. During their lifetime trees will experience great changes in their environment, both climatic and social, and these changes occur continuously.
Tree Responses
Tree physiology occurs on a cellular level. Tree physiology is the ability of trees to sense the environment and adjust accordingly. Tree response strategies depend on quick and reliable recognition systems. Examples of stimuli which trees perceive and can react to include the reaction responses listed below.
Light
In trees, blue light wave lengths can be used to cue developmental signals. Many tree organs contain photo-sensitive compounds (phototropins, cryptochromes, phytochromes, and the hormone auxin) each reacting very specifically to blue wave lengths of light. These light sensors tell the tree if it's day or night, how long the day is, how much light is available and from where the light is coming. Leaves turn direction to maximize sunlight exposure. Shoots grow towards light and roots usually grow away from light. They are directed by the photo-sensitive compounds mentioned above. Many trees exhibit certain phenomena at specific times of the day, for example certain flowers open only in the mornings.
Trees keep track of the time of the day with a molecular clock. This internal clock is set to the solar clock every day using sunlight. The internal clock coupled with the ability to perceive light allows trees to measure the time of the day and also find the season of the year. This is how many trees know when to flower or change leaf color in autumn. The seeds of many trees sprout only after they are exposed to light. Trees are also able to sense the quality of light and respond appropriately, for example in low light conditions trees produce more photosynthetic pigments whereas when the light is very bright and/or if the levels of harmful ultra-violet light increase, trees produce more of their protective pigments that act as sunscreens.
Temperature
In response to extremes of temperature trees can produce various proteins. As temperatures fall, production of antifreeze proteins and dehydrins increase. These protect the tree from the damaging effects of ice formation and falling rates of enzyme catalysis at low temperatures. As temperatures rise, production of heat shock proteins increases along with increased photo-respiration at high temperatures. Metabolic imbalances associated with temperature extremes result in the build-up of reactive oxygen species, which can be countered by antioxidant systems.
Cell membranes are also affected by changes in temperature and can cause the membrane to lose its fluid properties and become a gel in cold conditions or to become leaky in hot conditions. This can affect the movement of compounds across the membrane. To prevent these changes, trees can change the composition of their membranes. In cold conditions more unsaturated fatty acids are placed in the membrane and in hot conditions more saturated fatty acids are inserted.
Trees can avoid overheating by minimizing the amount of sunlight absorbed and by enhancing the cooling effects of wind and transpiration. Trees can reduce light absorption using reflective leaf hairs, scales, and waxes. These features are so common in warm dry regions that these habitats can be seen to form a ‘silvery landscape’ as the light scatters off the canopies. Some species can move their leaves throughout the day so that they are always orientated to avoid the sun on hot days (paraheliotropism). Knowledge of these mechanisms has been important for breeding heat stress tolerance in new cultivars.
Also in warmer weather, terpenes are released into the atmosphere surrounding conifer forests on hot summer days, where they act as a natural form of cloud seeding. Scientists now realize that the resulting clouds that form, help to block sunlight and cool the forest, allowing the forest to regulate its temperature. This function is similar to the way people sweat when they become overheated and the evaporating moisture on the skin, cools the body down.
Water
The ability of trees to access water depends on the structure of their roots and on the water potential of the root cells. If there is too little water then tissues will dehydrate and the tree may die. In very dry soil, trees close their stomata to reduce transpiration and prevent water loss. The closing of the stomata is often mediated by chemical signals from the root (i.e. abscisic acid). If the drought continues, the tree tissues will dehydrate resulting in a loss of turgor pressure that is visible as wilting. As well as closing their stomata, most trees can also respond to drought by altering their water potential (osmotic adjustment) and increasing root growth. The response to drought also causes vibrations in the trunk when the flow of water from the roots to the leaves is interrupted. The vibrations are heard at ultrasonic levels and may be considered a scream or an alarm to other trees nearby. Trees that are adapted to dry environments have a range of more specialized mechanisms to maintain water and/or protect tissues when desiccation occurs. This remarkable mechanism allows trees to lift water as high as 400 feet (120 m) by harnessing the gradient created by transpiration from the leaves.
Too much water can also damage trees. If the soil becomes waterlogged then the soil will become anoxic (low in oxygen) which can kill the roots of the tree within days. Trees can’t avoid waterlogging but many species overcome the lack of oxygen in the soil by transporting oxygen to the root from tissues that are not submerged. Species that are tolerant of waterlogging develop specialized roots near the soil surface and parenchyma cells to allow the diffusion of oxygen from the shoot to the root. Roots that are not killed outright may also switch to less oxygen-hungry forms of cellular respiration. Species that are frequently submerged have evolved more elaborate mechanisms that maintain root oxygen levels, perhaps most notable are the dramatic aerial roots seen in mangrove and cypress forests.
Carbon Dioxide (CO2) Concentration
Carbon dioxide is vital for tree growth as it is the substrate for photosynthesis. Trees take in carbon dioxide through stomatal pores on their leaves. At the same time as carbon dioxide enters the stomata, moisture escapes. This trade-off between carbon dioxide gain and water loss is central to tree productivity. The trade-off is all the more critical as the enzyme used to capture carbon dioxide is only efficient when there is a high concentration of carbon dioxide in the leaf. Some trees overcome this difficulty by concentrating carbon dioxide within their leaves using C4 carbon fixation. However, most species use C3 carbon fixation and must open their stomata to take in CO2 whenever photosynthesis is taking place.
The concentration of carbon dioxide in the atmosphere is rising due to deforestation, global warming, and the combustion of fossil fuels. This would be expected to increase the efficiency of photosynthesis and possibly increase the overall rate of tree growth. This possibility has attracted considerable interest from researchers in recent years, as an increased rate of tree growth could absorb some of the excess carbon dioxide and reduce the rate of global warming. Unfortunately, detrimental impacts of global warming, such as increased instances of heat and drought stress, mean that the overall effect is likely to be a reduction in tree productivity. Furthermore, reduced tree productivity would be expected to accelerate the rate of global warming. Overall, these observations point to the importance of avoiding further increases in atmospheric carbon dioxide rather than risking runaway climate change.
Wind
The main impact of wind on trees is through its influence on the canopy, which in turn influences the way leaves regulate moisture, heat, and carbon dioxide. When no wind is present, a layer of still air builds up around each leaf. This is known as the boundary layer and effectively insulates the leaf from the environment, providing an atmosphere rich in moisture and less prone to convective heating or cooling. As wind speed increases, the leaf environment becomes more closely linked to the surrounding environment. It may become difficult for the tree to retain moisture as it is exposed to dry air. On the other hand, a moderately high wind allows the tree to cool its leaves more easily when exposed to full sunlight. Trees are not entirely passive in their interaction with wind. Trees can make their leaves less vulnerable to changes in wind speed, by coating their leaves in fine hairs (trichomes) to break up the air flow and increase the boundary layer. In fact, leaf and canopy dimensions are often finely controlled to manipulate the boundary layer depending on the prevailing environmental conditions.
In areas where very strong winds are common, trees respond by reducing their above ground growth (known as dwarfing) and by strengthening their stems. Trees have a particularly well developed capacity to reinforce their trunks when exposed to wind In a cross section view of the trunk, it will have an oval shape with the longer dimension parallel to the prevailing wind.
In the most extreme cases, trees can be mortally damaged or uprooted by wind. This is a particular problem in hurricane-prone regions. When this type of disturbance occurs in natural systems, the only solution is to ensure that there is an adequate stock of seeds or seedlings to quickly take the place of the mature trees that have been lost, although in many cases a successional stage will be needed before the ecosystem can be restored to its former state.
Electrophysiology
Tree cells can be electrically excited and can display rapid electrical responses to environmental stimuli. These action potentials can influence processes such as tree movements, wound responses, respiration rate, photosynthesis, and flowering. The electrical responses can cause the synthesis of numerous organic molecules and accomplish behavioral responses to environmental, communicative, and ecological contexts. Cells in nearly all living organisms can be electrically excitable.
Cooperative Protection
When a tree is wounded or attacked by a native pest or disease, the tree will release a chemical odor, which can be noticed if you were to crush or tear a leaf. This odor serves as a volatile signal or an alarm signal that neighboring trees detect and prepare for the attack by producing chemicals that defend the tree from the insect or disease.
Trees have many strategies to fight off pests. For example, they can produce different toxins against invaders or they can induce a rapid cell death to kill diseased cells that hinders the disease from spreading throughout the tree. Another type of response is to attract predators, such as birds, that will eat an insect pest. This signal process has also been detected in the forest as an interlocking root system, assisted by mycorrhizae. Trees cannot always respond this well to exotic pests or diseases such as Dutch Elm Disease.
Response Summary
A tree's reactive behavior is mediated by phytochromes, kinins, hormones, antibiotic or other chemical releases, changes of water and chemical transport, and several other less frequent means. These responses are generally slow, taking at minimum a number of hours to accomplish, and can best be observed with time-lapse cinematography, but rapid movements can occur as well.
A tree's survival has developed from growing and evolving on this planet for millions of years. All of the reactions mentioned above have been observed and studied by researchers for many years.
Tree physiology occurs on a cellular level. Tree physiology is the ability of trees to sense the environment and adjust accordingly. Tree response strategies depend on quick and reliable recognition systems. Examples of stimuli which trees perceive and can react to include the reaction responses listed below.
Light
In trees, blue light wave lengths can be used to cue developmental signals. Many tree organs contain photo-sensitive compounds (phototropins, cryptochromes, phytochromes, and the hormone auxin) each reacting very specifically to blue wave lengths of light. These light sensors tell the tree if it's day or night, how long the day is, how much light is available and from where the light is coming. Leaves turn direction to maximize sunlight exposure. Shoots grow towards light and roots usually grow away from light. They are directed by the photo-sensitive compounds mentioned above. Many trees exhibit certain phenomena at specific times of the day, for example certain flowers open only in the mornings.
Trees keep track of the time of the day with a molecular clock. This internal clock is set to the solar clock every day using sunlight. The internal clock coupled with the ability to perceive light allows trees to measure the time of the day and also find the season of the year. This is how many trees know when to flower or change leaf color in autumn. The seeds of many trees sprout only after they are exposed to light. Trees are also able to sense the quality of light and respond appropriately, for example in low light conditions trees produce more photosynthetic pigments whereas when the light is very bright and/or if the levels of harmful ultra-violet light increase, trees produce more of their protective pigments that act as sunscreens.
Temperature
In response to extremes of temperature trees can produce various proteins. As temperatures fall, production of antifreeze proteins and dehydrins increase. These protect the tree from the damaging effects of ice formation and falling rates of enzyme catalysis at low temperatures. As temperatures rise, production of heat shock proteins increases along with increased photo-respiration at high temperatures. Metabolic imbalances associated with temperature extremes result in the build-up of reactive oxygen species, which can be countered by antioxidant systems.
Cell membranes are also affected by changes in temperature and can cause the membrane to lose its fluid properties and become a gel in cold conditions or to become leaky in hot conditions. This can affect the movement of compounds across the membrane. To prevent these changes, trees can change the composition of their membranes. In cold conditions more unsaturated fatty acids are placed in the membrane and in hot conditions more saturated fatty acids are inserted.
Trees can avoid overheating by minimizing the amount of sunlight absorbed and by enhancing the cooling effects of wind and transpiration. Trees can reduce light absorption using reflective leaf hairs, scales, and waxes. These features are so common in warm dry regions that these habitats can be seen to form a ‘silvery landscape’ as the light scatters off the canopies. Some species can move their leaves throughout the day so that they are always orientated to avoid the sun on hot days (paraheliotropism). Knowledge of these mechanisms has been important for breeding heat stress tolerance in new cultivars.
Also in warmer weather, terpenes are released into the atmosphere surrounding conifer forests on hot summer days, where they act as a natural form of cloud seeding. Scientists now realize that the resulting clouds that form, help to block sunlight and cool the forest, allowing the forest to regulate its temperature. This function is similar to the way people sweat when they become overheated and the evaporating moisture on the skin, cools the body down.
Water
The ability of trees to access water depends on the structure of their roots and on the water potential of the root cells. If there is too little water then tissues will dehydrate and the tree may die. In very dry soil, trees close their stomata to reduce transpiration and prevent water loss. The closing of the stomata is often mediated by chemical signals from the root (i.e. abscisic acid). If the drought continues, the tree tissues will dehydrate resulting in a loss of turgor pressure that is visible as wilting. As well as closing their stomata, most trees can also respond to drought by altering their water potential (osmotic adjustment) and increasing root growth. The response to drought also causes vibrations in the trunk when the flow of water from the roots to the leaves is interrupted. The vibrations are heard at ultrasonic levels and may be considered a scream or an alarm to other trees nearby. Trees that are adapted to dry environments have a range of more specialized mechanisms to maintain water and/or protect tissues when desiccation occurs. This remarkable mechanism allows trees to lift water as high as 400 feet (120 m) by harnessing the gradient created by transpiration from the leaves.
Too much water can also damage trees. If the soil becomes waterlogged then the soil will become anoxic (low in oxygen) which can kill the roots of the tree within days. Trees can’t avoid waterlogging but many species overcome the lack of oxygen in the soil by transporting oxygen to the root from tissues that are not submerged. Species that are tolerant of waterlogging develop specialized roots near the soil surface and parenchyma cells to allow the diffusion of oxygen from the shoot to the root. Roots that are not killed outright may also switch to less oxygen-hungry forms of cellular respiration. Species that are frequently submerged have evolved more elaborate mechanisms that maintain root oxygen levels, perhaps most notable are the dramatic aerial roots seen in mangrove and cypress forests.
Carbon Dioxide (CO2) Concentration
Carbon dioxide is vital for tree growth as it is the substrate for photosynthesis. Trees take in carbon dioxide through stomatal pores on their leaves. At the same time as carbon dioxide enters the stomata, moisture escapes. This trade-off between carbon dioxide gain and water loss is central to tree productivity. The trade-off is all the more critical as the enzyme used to capture carbon dioxide is only efficient when there is a high concentration of carbon dioxide in the leaf. Some trees overcome this difficulty by concentrating carbon dioxide within their leaves using C4 carbon fixation. However, most species use C3 carbon fixation and must open their stomata to take in CO2 whenever photosynthesis is taking place.
The concentration of carbon dioxide in the atmosphere is rising due to deforestation, global warming, and the combustion of fossil fuels. This would be expected to increase the efficiency of photosynthesis and possibly increase the overall rate of tree growth. This possibility has attracted considerable interest from researchers in recent years, as an increased rate of tree growth could absorb some of the excess carbon dioxide and reduce the rate of global warming. Unfortunately, detrimental impacts of global warming, such as increased instances of heat and drought stress, mean that the overall effect is likely to be a reduction in tree productivity. Furthermore, reduced tree productivity would be expected to accelerate the rate of global warming. Overall, these observations point to the importance of avoiding further increases in atmospheric carbon dioxide rather than risking runaway climate change.
Wind
The main impact of wind on trees is through its influence on the canopy, which in turn influences the way leaves regulate moisture, heat, and carbon dioxide. When no wind is present, a layer of still air builds up around each leaf. This is known as the boundary layer and effectively insulates the leaf from the environment, providing an atmosphere rich in moisture and less prone to convective heating or cooling. As wind speed increases, the leaf environment becomes more closely linked to the surrounding environment. It may become difficult for the tree to retain moisture as it is exposed to dry air. On the other hand, a moderately high wind allows the tree to cool its leaves more easily when exposed to full sunlight. Trees are not entirely passive in their interaction with wind. Trees can make their leaves less vulnerable to changes in wind speed, by coating their leaves in fine hairs (trichomes) to break up the air flow and increase the boundary layer. In fact, leaf and canopy dimensions are often finely controlled to manipulate the boundary layer depending on the prevailing environmental conditions.
In areas where very strong winds are common, trees respond by reducing their above ground growth (known as dwarfing) and by strengthening their stems. Trees have a particularly well developed capacity to reinforce their trunks when exposed to wind In a cross section view of the trunk, it will have an oval shape with the longer dimension parallel to the prevailing wind.
In the most extreme cases, trees can be mortally damaged or uprooted by wind. This is a particular problem in hurricane-prone regions. When this type of disturbance occurs in natural systems, the only solution is to ensure that there is an adequate stock of seeds or seedlings to quickly take the place of the mature trees that have been lost, although in many cases a successional stage will be needed before the ecosystem can be restored to its former state.
Electrophysiology
Tree cells can be electrically excited and can display rapid electrical responses to environmental stimuli. These action potentials can influence processes such as tree movements, wound responses, respiration rate, photosynthesis, and flowering. The electrical responses can cause the synthesis of numerous organic molecules and accomplish behavioral responses to environmental, communicative, and ecological contexts. Cells in nearly all living organisms can be electrically excitable.
Cooperative Protection
When a tree is wounded or attacked by a native pest or disease, the tree will release a chemical odor, which can be noticed if you were to crush or tear a leaf. This odor serves as a volatile signal or an alarm signal that neighboring trees detect and prepare for the attack by producing chemicals that defend the tree from the insect or disease.
Trees have many strategies to fight off pests. For example, they can produce different toxins against invaders or they can induce a rapid cell death to kill diseased cells that hinders the disease from spreading throughout the tree. Another type of response is to attract predators, such as birds, that will eat an insect pest. This signal process has also been detected in the forest as an interlocking root system, assisted by mycorrhizae. Trees cannot always respond this well to exotic pests or diseases such as Dutch Elm Disease.
Response Summary
A tree's reactive behavior is mediated by phytochromes, kinins, hormones, antibiotic or other chemical releases, changes of water and chemical transport, and several other less frequent means. These responses are generally slow, taking at minimum a number of hours to accomplish, and can best be observed with time-lapse cinematography, but rapid movements can occur as well.
A tree's survival has developed from growing and evolving on this planet for millions of years. All of the reactions mentioned above have been observed and studied by researchers for many years.
Tree Movements
Trees appear immobile because they are rooted in one place. However, time lapse photography reveals that parts of trees move frequently. Trees move in response to several environmental stimuli such as: light, gravity and mechanical disturbances. These movements fall into two groups: tropisms and nastic movements.
Tropism
A tropism is a plant movement that is determined by the direction of a biological stimulus. Movement toward an environmental stimulus is called a positive tropism, and movement away from a stimulus is called a negative tropism. Each kind of tropism is named for its stimulus. For example, a plant movement in response to light coming from one particular direction is called a phototropism.
Gravitropism
Gravitropism is a plant growth response to gravity. A root usually grows downward and a stem usually grows upward. Like phototropism, gravitropism appears to be regulated by auxins. It occurs when a seedling is placed horizontally, auxins accumulate along the lower sides of the root and the stem. This concentration of auxins stimulates cell elongation along the lower side of the stem, and the stem grows upward. A similar concentration of auxins inhibits cell elongation on the lower side of the root, and thus the root grows downward.
Chemotropism
Chemotropism is a plant growth response to a chemical. For example, after a flower is pollinated, a pollen tube grows down through the stigma and style and enters the ovule through the micropyle to produce a seed.
Nastic Movements
Some plants make nastic movements. Nastic movements differ from tropic movements in that the direction of tropic responses depends on the direction of the stimulus, whereas the direction of nastic movements is independent of the stimulus's position. Nastic movements are non-directional responses to stimuli (e.g. temperature, humidity, light), and are usually associated with plants. The movement can be due to changes in turgor or changes in growth. The rate or frequency of these responses increases as intensity of the stimulus increases. An example of such a response is the opening and closing of flowers. This movement is caused by the rapid loss of turgor pressure (water pressure) in certain cells, a process similar to that which occurs in guard cells in order to close stomata. Physical stimulation of the plant leaf causes potassium ions to be pumped out of the cells at the base of the leaflets and petioles. Water then moves out of the cells by osmosis. As the cells shrink, the plant leaves move. It is believed that the folding of a plant's leaves in response to touch is to discourage insect feeding.
Photoperiodism
Many angiosperms flower at about the same time every year. This occurs even though they may have started growing at different times. Their flowering is a response to the changing length of day and night as the season progresses. The phenomenon is called photoperiodism. It also helps promote cross pollination.
Photoperiodism is the mechanism by which many of our hardwood trees regulate their various life processes, including preparing for winter. Many plants have the ability to adjust their cycles to the changes in the amount of available daylight.
The day length at a latitude of North Carolina on the longest day of summer is approximately 14 hours and 20 minutes. By the autumn equinox, the length of day and night will be approximately equal to each other. At the winter solstice, the day length is about 9 hours and 45 minutes. At the onset of leaf color change, towards the end of September, the day length is decreasing at a rate of about 2 minutes per day.
Nyctinastic Movements
Nyctinastic movements are plant movements in response to the daily cycle of light and dark. Examples of trees that demonstrate these movements include honeylocust and silk trees. Both trees move their leaf blades horizontally during daylight hours and vertically at night. The nyctinastic movements involve the osmotic mechanism, but the changes in turgor pressure are more gradual.
Trees appear immobile because they are rooted in one place. However, time lapse photography reveals that parts of trees move frequently. Trees move in response to several environmental stimuli such as: light, gravity and mechanical disturbances. These movements fall into two groups: tropisms and nastic movements.
Tropism
A tropism is a plant movement that is determined by the direction of a biological stimulus. Movement toward an environmental stimulus is called a positive tropism, and movement away from a stimulus is called a negative tropism. Each kind of tropism is named for its stimulus. For example, a plant movement in response to light coming from one particular direction is called a phototropism.
Gravitropism
Gravitropism is a plant growth response to gravity. A root usually grows downward and a stem usually grows upward. Like phototropism, gravitropism appears to be regulated by auxins. It occurs when a seedling is placed horizontally, auxins accumulate along the lower sides of the root and the stem. This concentration of auxins stimulates cell elongation along the lower side of the stem, and the stem grows upward. A similar concentration of auxins inhibits cell elongation on the lower side of the root, and thus the root grows downward.
Chemotropism
Chemotropism is a plant growth response to a chemical. For example, after a flower is pollinated, a pollen tube grows down through the stigma and style and enters the ovule through the micropyle to produce a seed.
Nastic Movements
Some plants make nastic movements. Nastic movements differ from tropic movements in that the direction of tropic responses depends on the direction of the stimulus, whereas the direction of nastic movements is independent of the stimulus's position. Nastic movements are non-directional responses to stimuli (e.g. temperature, humidity, light), and are usually associated with plants. The movement can be due to changes in turgor or changes in growth. The rate or frequency of these responses increases as intensity of the stimulus increases. An example of such a response is the opening and closing of flowers. This movement is caused by the rapid loss of turgor pressure (water pressure) in certain cells, a process similar to that which occurs in guard cells in order to close stomata. Physical stimulation of the plant leaf causes potassium ions to be pumped out of the cells at the base of the leaflets and petioles. Water then moves out of the cells by osmosis. As the cells shrink, the plant leaves move. It is believed that the folding of a plant's leaves in response to touch is to discourage insect feeding.
Photoperiodism
Many angiosperms flower at about the same time every year. This occurs even though they may have started growing at different times. Their flowering is a response to the changing length of day and night as the season progresses. The phenomenon is called photoperiodism. It also helps promote cross pollination.
Photoperiodism is the mechanism by which many of our hardwood trees regulate their various life processes, including preparing for winter. Many plants have the ability to adjust their cycles to the changes in the amount of available daylight.
The day length at a latitude of North Carolina on the longest day of summer is approximately 14 hours and 20 minutes. By the autumn equinox, the length of day and night will be approximately equal to each other. At the winter solstice, the day length is about 9 hours and 45 minutes. At the onset of leaf color change, towards the end of September, the day length is decreasing at a rate of about 2 minutes per day.
Nyctinastic Movements
Nyctinastic movements are plant movements in response to the daily cycle of light and dark. Examples of trees that demonstrate these movements include honeylocust and silk trees. Both trees move their leaf blades horizontally during daylight hours and vertically at night. The nyctinastic movements involve the osmotic mechanism, but the changes in turgor pressure are more gradual.
Seasonal Responses
In non-tropical areas, tree responses are strongly influenced by seasonal changes. For example, many trees shed their leaves in the fall, and most trees flower only at certain times of the year. Trees are able to sense seasonal changes. Although temperature changes are involved in some case and to certain degrees, trees mark the seasons primarily by sensing changes in night length.
Senescence
Why do leaves turn color in the autumn? Senescence is the process that occurs when chlorophyll production drops off as day length decreases and the tree prepares for dormancy. As the photoperiod decreases, the plants ability to synthesize chlorophyll becomes reduced. The green part of the light spectrum is no longer reflected and other compounds, chemicals called "anthocyanins" (reds), “xanthophyll’s” (oranges), and "carotenoids" (yellows), become the dominant pigments in the leaves. The leaves are changed and everything but the cell walls and nutrient depleted protoplasm are gradually drawn into the stems and roots in a very efficient manner. Additionally, senescing cells in oaks contain high quantities of tannins in the leaves which are responsible for brown colors.
The intensity or brilliance of the color change is influenced by weather conditions during the period of declining chlorophyll production. Moist soils following a good growing season contribute to better displays of vibrant colors. A series of sunny days and cool nights that remain above freezing will also result in a more colorful display. On the other hand, the warm days of autumn weather will generally reduce the color quality.
Dormancy
Dormancy is a period in a plant's life of decreased metabolism. In the deciduous hardwoods of the temperate regions, this period is usually referred to by the non-technical term "winter". In preparation for winter and to prevent or minimize damage from the cold, plant cells switch from production of chlorophyll for growth, to production of sugars and amino acids, which act as antifreeze for the plant. As already mentioned, chemicals in the leaves are drawn into the stems and eventually to the roots during this process.
The cold temperatures prevent trees and plants from functioning in at least three ways. Water would freeze in the plant tissues, causing cell rupture. Water in the upper soil layers often freezes, making absorption impossible. Finally, the low temperatures are far below what is necessary for the proper function of the enzymes that control a tree’s metabolic processes, such as photosynthesis and respiration. To avoid these environmental limitations, trees prepare for dormancy in the autumn.
At the base of the leaf petiole, where the leaf is attached to the stem, two types of cells are formed:
In non-tropical areas, tree responses are strongly influenced by seasonal changes. For example, many trees shed their leaves in the fall, and most trees flower only at certain times of the year. Trees are able to sense seasonal changes. Although temperature changes are involved in some case and to certain degrees, trees mark the seasons primarily by sensing changes in night length.
Senescence
Why do leaves turn color in the autumn? Senescence is the process that occurs when chlorophyll production drops off as day length decreases and the tree prepares for dormancy. As the photoperiod decreases, the plants ability to synthesize chlorophyll becomes reduced. The green part of the light spectrum is no longer reflected and other compounds, chemicals called "anthocyanins" (reds), “xanthophyll’s” (oranges), and "carotenoids" (yellows), become the dominant pigments in the leaves. The leaves are changed and everything but the cell walls and nutrient depleted protoplasm are gradually drawn into the stems and roots in a very efficient manner. Additionally, senescing cells in oaks contain high quantities of tannins in the leaves which are responsible for brown colors.
The intensity or brilliance of the color change is influenced by weather conditions during the period of declining chlorophyll production. Moist soils following a good growing season contribute to better displays of vibrant colors. A series of sunny days and cool nights that remain above freezing will also result in a more colorful display. On the other hand, the warm days of autumn weather will generally reduce the color quality.
Dormancy
Dormancy is a period in a plant's life of decreased metabolism. In the deciduous hardwoods of the temperate regions, this period is usually referred to by the non-technical term "winter". In preparation for winter and to prevent or minimize damage from the cold, plant cells switch from production of chlorophyll for growth, to production of sugars and amino acids, which act as antifreeze for the plant. As already mentioned, chemicals in the leaves are drawn into the stems and eventually to the roots during this process.
The cold temperatures prevent trees and plants from functioning in at least three ways. Water would freeze in the plant tissues, causing cell rupture. Water in the upper soil layers often freezes, making absorption impossible. Finally, the low temperatures are far below what is necessary for the proper function of the enzymes that control a tree’s metabolic processes, such as photosynthesis and respiration. To avoid these environmental limitations, trees prepare for dormancy in the autumn.
At the base of the leaf petiole, where the leaf is attached to the stem, two types of cells are formed:
- Parenchyma cells, which are very soft and are formed on the leaf side of the petiole.
- Suberized cells, which are waxy and impermeable, are formed on the tree side to act as a protective seal.
Eventually, the vascular bundles (the veins of the leaf), are the only things that hold the leaf to the tree. The vascular bundles eventually break, or are torn by the wind, and the leaf falls, leaving what is called a bundle scar, and a bud for next year’s growth
Most conifers that have different strategies to maintain their green parts during the winter and they have needles with a much different structure than broad leaves. However, in the case of conifers, the needles that have grown old after two to three years, no longer receive as much light and are shed each autumn.
Fallen Leaves
The dropping of tons of biomass per acre presents a tree losing significant amounts of valuable nutrients. Much of the sugars and valuable nutrients are resorbed from the leaves just before they separate from the tree, but the annual leaf drop still means the loss of a lot of good nutrients that will eventually decompose on the forest floor and be absorbed by the micronutrients that will release them back into the tree roots within the next couple of growing seasons. In the north temperate climates, dropped leaves become part of the "organic layer" on the surface of the soil, to be recycled in part, by the soil decomposers category of soil microorganisms.
Biostimulants
Biostimulants are products applied by humans to assist trees that are struggling in the urban environment. Biostimulants are living organisms or extracts from living things that can either help protect plants from disease or invigorate growth. They include powdered live forms of organic compounds such as mycorrhizal fungi (ecto- and endo-), beneficial fungi (Gliocladium, Tricoderma), and root-colonizing bacteria (Streptomycetes, Bacillus, Pseudomonas). They also include such things as yucca extracts (wetting agent), kelp, humates, protein hydrolysates, and glutens that provide slow release forms of nitrogen. Certain harmful soil pests can be controlled with chili pepper spray, garlic tea, and Neem oil (a broad-spectrum insecticide derived from the oil of Neem tree seeds).
The term 'biostimulant' has been used to describe various substances involved in horticultural production over the past few decades. They were originally developed to promote root growth in agriculture and increased root crops by 20 – 30%. They also were used to reduce stress, disease susceptibility, and improve root quality. In recent years, however, it has been established that a biostimulant is a substance that is not a plant nutrient or pesticide but in some manner has a positive impact on soil and plant health. The biostimulant may enhance metabolism and respiration, increase chlorophyll efficiency and production, increase antioxidants, nucleic acid synthesis, enhance nutrient availability and increase the water holding capacity of the soil.
The benefits of biostimulants have been recognized for some time and university research has been published showing various benefits to growing organisms. In several research studies, biostimulants have decreased certain problems leading to chlorosis by enabling the plant to absorb minerals not readily available within the soil profile. University research has demonstrated the effectiveness of biostimulants and how they help retain a healthy forest soil environment. Also biostimulants have been shown to enable antioxidant activity in both the soil and within the plant, especially when the plants are under stress. In many cases, biostimulants help the tree recover quickly from stress and rejuvenate the root system.
In the age where caution has to be utilized with chemicals placed on the landscape, biostimulants can help by utilizing organic means of replenishing nutrients into the soil, while enabling the plant to uptake products more efficiently. At the same time, biostimulants decrease the overall amount of nitrate rich fertilizers, pesticides, and fungicides that need to be applied. Biostimulants can also improve root growth after transplanting. However, the appropriate biostimulant must be used for the specific species, and many biostimulants do not meet the manufacturer's claims.
Most conifers that have different strategies to maintain their green parts during the winter and they have needles with a much different structure than broad leaves. However, in the case of conifers, the needles that have grown old after two to three years, no longer receive as much light and are shed each autumn.
Fallen Leaves
The dropping of tons of biomass per acre presents a tree losing significant amounts of valuable nutrients. Much of the sugars and valuable nutrients are resorbed from the leaves just before they separate from the tree, but the annual leaf drop still means the loss of a lot of good nutrients that will eventually decompose on the forest floor and be absorbed by the micronutrients that will release them back into the tree roots within the next couple of growing seasons. In the north temperate climates, dropped leaves become part of the "organic layer" on the surface of the soil, to be recycled in part, by the soil decomposers category of soil microorganisms.
Biostimulants
Biostimulants are products applied by humans to assist trees that are struggling in the urban environment. Biostimulants are living organisms or extracts from living things that can either help protect plants from disease or invigorate growth. They include powdered live forms of organic compounds such as mycorrhizal fungi (ecto- and endo-), beneficial fungi (Gliocladium, Tricoderma), and root-colonizing bacteria (Streptomycetes, Bacillus, Pseudomonas). They also include such things as yucca extracts (wetting agent), kelp, humates, protein hydrolysates, and glutens that provide slow release forms of nitrogen. Certain harmful soil pests can be controlled with chili pepper spray, garlic tea, and Neem oil (a broad-spectrum insecticide derived from the oil of Neem tree seeds).
The term 'biostimulant' has been used to describe various substances involved in horticultural production over the past few decades. They were originally developed to promote root growth in agriculture and increased root crops by 20 – 30%. They also were used to reduce stress, disease susceptibility, and improve root quality. In recent years, however, it has been established that a biostimulant is a substance that is not a plant nutrient or pesticide but in some manner has a positive impact on soil and plant health. The biostimulant may enhance metabolism and respiration, increase chlorophyll efficiency and production, increase antioxidants, nucleic acid synthesis, enhance nutrient availability and increase the water holding capacity of the soil.
The benefits of biostimulants have been recognized for some time and university research has been published showing various benefits to growing organisms. In several research studies, biostimulants have decreased certain problems leading to chlorosis by enabling the plant to absorb minerals not readily available within the soil profile. University research has demonstrated the effectiveness of biostimulants and how they help retain a healthy forest soil environment. Also biostimulants have been shown to enable antioxidant activity in both the soil and within the plant, especially when the plants are under stress. In many cases, biostimulants help the tree recover quickly from stress and rejuvenate the root system.
In the age where caution has to be utilized with chemicals placed on the landscape, biostimulants can help by utilizing organic means of replenishing nutrients into the soil, while enabling the plant to uptake products more efficiently. At the same time, biostimulants decrease the overall amount of nitrate rich fertilizers, pesticides, and fungicides that need to be applied. Biostimulants can also improve root growth after transplanting. However, the appropriate biostimulant must be used for the specific species, and many biostimulants do not meet the manufacturer's claims.
Tree Growth Regulators
All living organisms begin in the same form: as a single cell. That cell will divide and the resulting cells will continue dividing and differentiate into cells with various roles to carry out within the organism. This is the way human and animal life is on this planet and trees are no different. Plant growth can be determinate or indeterminate, meaning some plants will have a cycle of growth then a cessation of growth, breakdown of tissues, and then death (like a tomato plant) while others (like a ginkgo tree) will grow and remain active for hundreds of years. A tomato plant is fairly predictable and is said to have determinate growth, while the ginkgo tree has indeterminate growing potential. (Ginkgo trees have no genetic means of dying except by outside causes like a windstorm.) Development refers to the growth and differentiation of cells into tissues, organs and organ systems.
Tree Enzymes
Genetic information directs the synthesis and development of enzymes which are critical in all metabolic processes within the tree. Most enzymes are proteins in some form or another. They are produced in very minute quantities and are produced within the tree. Vitamins are vital in the activation of enzymes and are produced in the cytoplasm (a gel-like substance enclosed within the cell membrane) and the membranes of tree cells.
Tree Hormones
Genetic information also regulates the production of hormones. The major difference is that hormones are transported from one part of the plant to another as needed. In general, hormone and vitamin effects are similar and are difficult to distinguish in trees, and both are referred to in general as tree growth regulators.
The growth and development of a tree is influenced by genetic factors, external environmental factors, and chemical hormones inside the tree. Trees respond to many environmental factors such as light, gravity, water, inorganic nutrients, and temperature.
Tree hormones are chemical messengers that affect a tree's ability to respond to its environment. Hormones are organic compounds that are effective at very low concentrations. They are usually synthesized in one part of the plant and are transported to another location. They interact with specific target tissues to cause physiological responses, such as growth or fruit ripening. Each response is often the result of two or more hormones acting together. Because hormones stimulate or inhibit tree growth, many botanists also refer to them as tree growth regulators. In the Ginkgo tree, the protective chemicals fend off drought and diseases but then, unlike other trees, they remain in the ginkgo fighting diseases for as long as the tree lives. Many hormones can be synthesized in the laboratory, thereby increasing the quantity of hormones available for commercial applications.
Botanists recognize five major groups of hormones: auxins, gibberellins, ethylene, cytokines, and abscisic acid.
Auxins
Auxins are hormones involved in plant-cell elongation, apical dominance, and rooting. A well-known natural auxin is indoleacetic acid, or IAA which is produced in the apical meristem of the shoot. Developing seeds produce IAA, which stimulates the development of a fleshy fruit. For example, the removal of seeds from an apple prevents the fruit from enlarging. However, the application of IAA after removing the seeds causes the fruit to enlarge normally. Before a cell can elongate, the cell wall must become less rigid so that it can expand. IAA triggers an increase in the plasticity of cell walls, allowing this elongation to occur.
Synthetic Auxins
Chemists have synthesized several inexpensive compounds similar in structure to IAA. Synthetic auxins, like naphthalene acetic acid, of NAA, are used extensively to promote root formation on stem and leaf cuttings. When NAA is sprayed on young fruits of apple and olive trees, some of the fruits drop off so that the remaining fruits grow larger. When NAA is sprayed directly on maturing fruits, such as apples, pears and citrus fruits, several weeks before they are ready to be picked, NAA prevents the fruits from dropping off the trees before they are mature. The fact that auxins can have opposite effects, causing fruit to drop or preventing fruit from dropping, illustrates an important point. The effects of a hormone on a plant often depend on the stage of the plant's development.
NAA is also used to prevent the undesirable sprouting of stems from the base of ornamental trees. As previously discussed, stems contain a lateral bud at the base of each leaf. In many stems, these buds fail to sprout as long as the plant's shoot tip is still intact. The inhibition of lateral buds by the presence of the shoot tip is called apical dominance. If the shoot tip of a plant is removed, the lateral buds begin to grow. If IAA or NAA is applied to the cut tip of the stem, the lateral buds remain dormant. This adaptation is manipulated to cultivate beautiful ornamental trees.
Gibberellins
In the 1920's scientists in Japan discovered that a substance produced by the fungus Gibberella caused fungus-infected plants to grow abnormally tall. The substance, named gibberellin, was later found to be produced in small quantities by plants themselves. It has many effects on a plant, but primarily stimulates elongation growth. Spraying a plant with gibberellins will usually cause the plant to grow to a larger than expected height.
Almost all seedless grapes are sprayed with gibberellins to increase the size of the fruit and the distance between fruits on the stems. Beer makers use gibberellins to increase the alcohol content of beer by increasing the amount of sugar produced in the malting process. Gibberellins are also used to treat seeds of some food crops because they will break seed dormancy and promote uniform germination.
Ethylene
The hormone ethylene is responsible for the ripening of fruits. Unlike the other four classes of plant hormones, ethylene is a gas at room temperature. Ethylene gas diffuses easily through the air from one plant to another. The saying "One bad apple spoils the barrel" has its basis in the effects of ethylene gas. One rotting apple will produce ethylene gas, which stimulates nearby apples to ripen and eventually spoil because of over ripening.
Ethylene is usually applied in a solution of ethephon, a synthetic chemical that breaks down and releases ethylene gas. It is used to ripen bananas, honeydew melons and tomatoes. Oranges, lemons, and grapefruits often remain green when they are ripe. Although the fruit tastes good, consumers often will not buy them, because oranges are supposed to be orange. The application of ethylene to green citrus fruit causes the development of desirable citrus colors, such as orange and yellow. In some plant species, ethylene promotes abscission, which is the detachment of leaves, flowers, or fruits from a plant. Cherries and walnuts are harvested with mechanical tree shakers. Ethylene treatment increases the number of fruits that fall to the ground when the trees are shaken. Leaf abscission also has an adaptive advantage for the plant. Dead, damaged or infected leaves drop to the ground rather than shading healthy leaves or spreading disease.
Cytokines
Cytokines promote cell division in plants. Produced in the developing shoots, roots, fruits, and seeds of a plant, cytokines are very important in the culturing of plant tissues in the laboratory. A high ratio of auxins to cytokines in a tissue-culture medium stimulates root formation. A low ratio promotes shoot formation. Cytokines are also used to promote lateral bud growth in flowering plants.
Abscisic Acid
Abscisic acid, or ABA, generally inhibits other hormones, such as the auxin IAA. It was originally thought to promote abscission, hence its name. Botanists now know that ethylene is the main abscission hormone. ABA helps to bring about dormancy in a plant's buds and maintains dormancy in its seeds. ABA causes the closure of a plant's stomata in response to drought. Water stressed leaves produce large amounts of ABA, which triggers potassium ions to be transported out of the guard cells. This causes stomata to close, and water is held in the leaf. It is too costly to synthesize ABA for commercial agriculture use.
Other Growth Regulators
Many growth regulators are widely used on ornamental plants. These substances do not fit into any of the five classes of hormones. For example, utility companies all over the country apply growth retardants to trees in order to prevent them from interfering with overhead utility lines. It is less expensive to apply these chemicals than to prune the trees, not to mention safer for the utility workers. Also, azalea growers sometimes apply a growth regulator to the terminal buds rather than hand-pruning them.
Conclusion
A tree's ability to dynamically adapt comes directly from their structure and specifically from the weaknesses that evolution has taught trees to build into their structure. They will continue to survive and reproduce long after mankind has left the planet.
All living organisms begin in the same form: as a single cell. That cell will divide and the resulting cells will continue dividing and differentiate into cells with various roles to carry out within the organism. This is the way human and animal life is on this planet and trees are no different. Plant growth can be determinate or indeterminate, meaning some plants will have a cycle of growth then a cessation of growth, breakdown of tissues, and then death (like a tomato plant) while others (like a ginkgo tree) will grow and remain active for hundreds of years. A tomato plant is fairly predictable and is said to have determinate growth, while the ginkgo tree has indeterminate growing potential. (Ginkgo trees have no genetic means of dying except by outside causes like a windstorm.) Development refers to the growth and differentiation of cells into tissues, organs and organ systems.
Tree Enzymes
Genetic information directs the synthesis and development of enzymes which are critical in all metabolic processes within the tree. Most enzymes are proteins in some form or another. They are produced in very minute quantities and are produced within the tree. Vitamins are vital in the activation of enzymes and are produced in the cytoplasm (a gel-like substance enclosed within the cell membrane) and the membranes of tree cells.
Tree Hormones
Genetic information also regulates the production of hormones. The major difference is that hormones are transported from one part of the plant to another as needed. In general, hormone and vitamin effects are similar and are difficult to distinguish in trees, and both are referred to in general as tree growth regulators.
The growth and development of a tree is influenced by genetic factors, external environmental factors, and chemical hormones inside the tree. Trees respond to many environmental factors such as light, gravity, water, inorganic nutrients, and temperature.
Tree hormones are chemical messengers that affect a tree's ability to respond to its environment. Hormones are organic compounds that are effective at very low concentrations. They are usually synthesized in one part of the plant and are transported to another location. They interact with specific target tissues to cause physiological responses, such as growth or fruit ripening. Each response is often the result of two or more hormones acting together. Because hormones stimulate or inhibit tree growth, many botanists also refer to them as tree growth regulators. In the Ginkgo tree, the protective chemicals fend off drought and diseases but then, unlike other trees, they remain in the ginkgo fighting diseases for as long as the tree lives. Many hormones can be synthesized in the laboratory, thereby increasing the quantity of hormones available for commercial applications.
Botanists recognize five major groups of hormones: auxins, gibberellins, ethylene, cytokines, and abscisic acid.
Auxins
Auxins are hormones involved in plant-cell elongation, apical dominance, and rooting. A well-known natural auxin is indoleacetic acid, or IAA which is produced in the apical meristem of the shoot. Developing seeds produce IAA, which stimulates the development of a fleshy fruit. For example, the removal of seeds from an apple prevents the fruit from enlarging. However, the application of IAA after removing the seeds causes the fruit to enlarge normally. Before a cell can elongate, the cell wall must become less rigid so that it can expand. IAA triggers an increase in the plasticity of cell walls, allowing this elongation to occur.
Synthetic Auxins
Chemists have synthesized several inexpensive compounds similar in structure to IAA. Synthetic auxins, like naphthalene acetic acid, of NAA, are used extensively to promote root formation on stem and leaf cuttings. When NAA is sprayed on young fruits of apple and olive trees, some of the fruits drop off so that the remaining fruits grow larger. When NAA is sprayed directly on maturing fruits, such as apples, pears and citrus fruits, several weeks before they are ready to be picked, NAA prevents the fruits from dropping off the trees before they are mature. The fact that auxins can have opposite effects, causing fruit to drop or preventing fruit from dropping, illustrates an important point. The effects of a hormone on a plant often depend on the stage of the plant's development.
NAA is also used to prevent the undesirable sprouting of stems from the base of ornamental trees. As previously discussed, stems contain a lateral bud at the base of each leaf. In many stems, these buds fail to sprout as long as the plant's shoot tip is still intact. The inhibition of lateral buds by the presence of the shoot tip is called apical dominance. If the shoot tip of a plant is removed, the lateral buds begin to grow. If IAA or NAA is applied to the cut tip of the stem, the lateral buds remain dormant. This adaptation is manipulated to cultivate beautiful ornamental trees.
Gibberellins
In the 1920's scientists in Japan discovered that a substance produced by the fungus Gibberella caused fungus-infected plants to grow abnormally tall. The substance, named gibberellin, was later found to be produced in small quantities by plants themselves. It has many effects on a plant, but primarily stimulates elongation growth. Spraying a plant with gibberellins will usually cause the plant to grow to a larger than expected height.
Almost all seedless grapes are sprayed with gibberellins to increase the size of the fruit and the distance between fruits on the stems. Beer makers use gibberellins to increase the alcohol content of beer by increasing the amount of sugar produced in the malting process. Gibberellins are also used to treat seeds of some food crops because they will break seed dormancy and promote uniform germination.
Ethylene
The hormone ethylene is responsible for the ripening of fruits. Unlike the other four classes of plant hormones, ethylene is a gas at room temperature. Ethylene gas diffuses easily through the air from one plant to another. The saying "One bad apple spoils the barrel" has its basis in the effects of ethylene gas. One rotting apple will produce ethylene gas, which stimulates nearby apples to ripen and eventually spoil because of over ripening.
Ethylene is usually applied in a solution of ethephon, a synthetic chemical that breaks down and releases ethylene gas. It is used to ripen bananas, honeydew melons and tomatoes. Oranges, lemons, and grapefruits often remain green when they are ripe. Although the fruit tastes good, consumers often will not buy them, because oranges are supposed to be orange. The application of ethylene to green citrus fruit causes the development of desirable citrus colors, such as orange and yellow. In some plant species, ethylene promotes abscission, which is the detachment of leaves, flowers, or fruits from a plant. Cherries and walnuts are harvested with mechanical tree shakers. Ethylene treatment increases the number of fruits that fall to the ground when the trees are shaken. Leaf abscission also has an adaptive advantage for the plant. Dead, damaged or infected leaves drop to the ground rather than shading healthy leaves or spreading disease.
Cytokines
Cytokines promote cell division in plants. Produced in the developing shoots, roots, fruits, and seeds of a plant, cytokines are very important in the culturing of plant tissues in the laboratory. A high ratio of auxins to cytokines in a tissue-culture medium stimulates root formation. A low ratio promotes shoot formation. Cytokines are also used to promote lateral bud growth in flowering plants.
Abscisic Acid
Abscisic acid, or ABA, generally inhibits other hormones, such as the auxin IAA. It was originally thought to promote abscission, hence its name. Botanists now know that ethylene is the main abscission hormone. ABA helps to bring about dormancy in a plant's buds and maintains dormancy in its seeds. ABA causes the closure of a plant's stomata in response to drought. Water stressed leaves produce large amounts of ABA, which triggers potassium ions to be transported out of the guard cells. This causes stomata to close, and water is held in the leaf. It is too costly to synthesize ABA for commercial agriculture use.
Other Growth Regulators
Many growth regulators are widely used on ornamental plants. These substances do not fit into any of the five classes of hormones. For example, utility companies all over the country apply growth retardants to trees in order to prevent them from interfering with overhead utility lines. It is less expensive to apply these chemicals than to prune the trees, not to mention safer for the utility workers. Also, azalea growers sometimes apply a growth regulator to the terminal buds rather than hand-pruning them.
Conclusion
A tree's ability to dynamically adapt comes directly from their structure and specifically from the weaknesses that evolution has taught trees to build into their structure. They will continue to survive and reproduce long after mankind has left the planet.
- Sources:
- Andersen C.P., “Ozone stress and changes below ground: limiting root and soil processes”, Phyton, Special issue “Root-soil interactions”, Vol. 40, 1-4:7-12, 2000.
- Ake Strid and Robert J. Porra, "Alterations in Pigment Content in Leaves of Pisum sativum After Exposure to Supplementary UV-B", Plant and Cell Physiology, Vol. 33, No. 7, 1992.
- Bassirirad H., “Kinetics of nutrient uptake by roots: responses to global change”, New Phytologist, 147: 155-169, 2000.
- Battisti A. Forest, “Climate change - lessons from insects”, Forest@, 1(1):17-24, on-line journal published by the Italian Society of Silviculture and Forest Ecology, 2004.
- Bisgrove R. and P. Hadley, “Gardening in the Global Greenhouse: the Impacts of Climate Change on Gardens in the UK”, Technical Report, November 2002.
- Brenner, E., Stahlberg, R., Mancuso, S., Vivanco, J., Baluska, F., Vanvolkenburgh, E., "Plant Neurobiology: an integrated view of plant signaling", Trends in Plant Science, 2006.
- Coder, Dr. Kim D. "Tree Root Growth Requirements", City Trees, The Journal of The Society of Municipal Arborists, Vol 38, Number 2 March/April 2002.
- Cregg B.M., Dix M.E., “Tree moisture stress and insect damage in urban areas in relation to heat island effects”, Journal of Arboriculture, 27: 8-17, 2001.
- Ferrini, F., A. Fini, G. Amoroso, P. Frangi, “Mulching of ornamental trees: effects on growth and physiology”. Arboriculture and Urban Forestry, 34(3): 15-162, 2008.
- Jim, C.J., “Green-space preservation and allocation for sustainable greening of compact cities”, Cities, 21(4): 311-320, 2004.
- Lloyd -Jones, David, “Tree Morphogenesis Book 1 Reduction via Thinning”, Kindle Edition, 2013.
- Phillips, Leonard, "Root Physiology", City Trees, The Journal of The Society of Municipal Arborists Vol 35, Number 4 July/August 1999: p. 13-16.
- Pittenger, Dennis R., California Master Gardener Handbook, University of California, Division of Agriculture and Natural Resources, ANR Publications, 2002.
- Richardson, Rosemary, Biology 203 Course, Bellevue Community College Science Division, Spring 2003.
- Rindels, Sherry, "Tree Root Systems", ISU Extension, Prepared by Department of Horticulture Iowa State University, Ames, Iowa, pp. 43-44, April 1, 1992 issue.
- Smiley, T., S. Lilly, P. Kelsey, “Fertilizing trees and shrubs. Determining if, when, and what to use”, Arborist News 2:17-21, 2002.
- Way, D.A. "Tree phenology responses to warming: spring forward, fall back?" Tree Physiology, 31:469-471, 2002.
- Xiong, L., Zhu, J. K., "Abiotic Stress Signal Transduction in Plants: Molecular and Genetic Perspectives", Physiologia Plantarum, 12 (2):152, 2001.
The test that follows contains 80 questions. Before taking the test be sure you have read the article carefully. The passing grade is 80% on the entire test.
LA CES will award 6.0 PDH (HSW) credits for a passing grade. This course has also been approved by North Carolina Board of LA and New Jersey Board of Architects for 6.0 credits.
The cost for taking this test is $20 per credit. If you purchase an annual subscription for 12 credits, the cost per credit is reduced by 50% (see Annual Subscription link below). We will report your passing test score to LA CES. If you are also ISA* certified we will report your passing score to ISA for no additional cost. Please be sure to add your ISA Cert. number when you sign in. Tests with passing scores may be submitted only once to each organization.
*ISA has approved this course for 4.0 CEUs which may be applied toward Certified Arborist, Tree Worker Specialist, Municipal Specialist, or BCMA science credits.
To take the test by the pay per test option, click on the 'Pay Now' button below where you can send payment online securely with your credit card or Pay Pal account. After your payment is submitted, click on ‘Return to merchant / gibneyCE.com’. That will take you to the test sign in page followed by the test. If you are an ISA and/or CLARB member, please be sure to include your certification/member number(s) along with your LA license and ASLA numbers.
To take the test as an annual subscriber with reduced rates, click on Password and enter your test password which will take you to the test sign in page. If you would like to become a subscriber see our Annual Subscription page for details.
When you have finished answering all questions you will be prompted to click ‘next’ to send your answers to gibneyCE.com. You can then click ‘next’ to view your test summary. A test review of your answers is available upon request. You can spend as much time as you would like to take the test but it is important not to leave the test site until you have answered all the questions and see the 'sending your answers' response.
Test re-takes are allowed, however you will have to pay for the retake if you are using the pay per test option.
All passing test scores are sent from gibneyCE.com to your organization(s) at the end of every month and they will appear on your certification record 4 to 6 weeks after that.
LA CES maintains a record of earned PDH credits on their website http://laces.asla.org/
ISA maintains a record of earned CEU credits on their website http://www.isa-arbor.com/