#1 Read About Root Development
Edited by Len Phillips, updated January 2023
Sections You may go directly to the section by clicking on titles listed here.
Edited by Len Phillips, updated January 2023
Sections You may go directly to the section by clicking on titles listed here.
Note: Click on green text in each section for more information.
Root Forms
Roots survive and grow where adequate water is available, temperatures are warm, and oxygen is present. Roots are limited by oxygen content, anaerobic conditions, and water depth. Near the base of the tree, deep-growing roots can be found because they are oxygenated through fissures and cracks in the soil generated as a result of mechanical forces moving the crown and trunk swaying from the wind.
Roots go through several forms:
The taproot - When a seed germinates, the primary root, or radicle as it is called in the embryo, emerges from the seed coat. The taproot system develops from the primary root. It is comprised of one major root that is thicker at its junction with the trunk or stem and tapers toward the growing tip. Taproots penetrate deep into the soil and are often used for food storage. They form small lateral roots and are often deep and provide excellent anchorage for the tree. With increasing soil density downward, taproot growth is restricted and lateral root growth increases. On fibrous root systems, most taproots disappear after the seedling stage. Taproot systems are found in many dicots and gymnosperms. One research report has indicated that less than 5% of all trees have a tap root system.
Fibrous root systems consist of many lateral roots that grow horizontally to stabilize a tree. Many if not all fibrous root systems develop from small adventitious roots that develop from stem tissue soon after germination, particularly in monocots. Fibrous roots are shallow, wide spreading, numerous, and good for soil erosion control because they cling to soil particles and hold the soil in place.
A combination of both root systems occurs in most species that grow a taproot as a seedling until a certain distance or obstacle such as a rock is encountered, and then the root system changes and continues to grow as a fibrous system. This frequently occurs with nursery grown trees when the tap root from the seedling encounters the edge of the container or is air-pruned at the bottom of the container.
Sloping Root Systems (also called heart root systems), have roots growing at many different angles from the horizontal to the vertical. Research shows that 15% of all trees have sloping root systems where the roots appear to radiate from a common point at the base of the trunk.
Aerial roots are roots that do not grow underground, but emerge from the trunk. There are a number of different types of aerial roots.
Propagation Roots – Many trees can produce "suckers" from roots. Suckers are adventitious woody shoots. Aspen groves, such as Pando in Utah's Fishlake National Forest, in which many trees are formed from one original tree, can cover large areas and all the trees in the grove grow from root suckers. All these trees are genetically identical.
Root Nodules – Some legumes and a few other plants produce root nodules, which contain bacteria or cyanobacteria that can "fix" nitrogen (make nitrogen into the nitrate form that is useful as a plant nutrient). When the bacteria enter the root, they induce the root to form "tumors" which are the nodules. The bacteria then grow and produce nitrogen forms in the nodules.
Root Systems
There are two basic types of roots, woody and non-woody.
Non-woody roots are found mostly in the upper few inches of soil. The primary function of these roots is to absorb water and nutrients. These are often called feeder roots. These roots also produce certain hormones (gibberelins and cytokinins) in the root meristem that are translocated through the tree to control growth and development. Non-woody roots have very little lignin in their cell walls. These roots live for a short time; from a few weeks to a year.
There are two types of structures that form on non-woody roots: root hairs and mycorrhizae.
1. Root hairs are organs that are extensions of epidermal cells. In some trees, particularly deciduous trees, the root hairs increase root surface area and increase nutrient and water uptake. Evergreen trees may not have root hairs but may possess mycorrhizae.
2. Mycorrhizae are organs composed of tree and fungus tissue, a composite structure. Mycorrhizae are a type of fungi that live on and in the feeder roots. These fungi do not cause any harm to the tree. In fact, for most species it is very beneficial for the tree to have this fungal association. Scientists have recently discovered that the tips of roots and mycorrhizae are involved in underground communication among trees and other plants.
Woody roots are large lateral roots that form at the flare or root collar (where the top of a root meets the bottom of the trunk). The primary purpose of woody roots are to support and anchor the tree. They also provide water and mineral transport as well as carbohydrate storage. These roots are distinct for each tree species and provide the framework for the tree's root system. The general direction for this framework is radial and horizontal and these roots are located 8 to 12 inches (20 – 30 cm) below the soil surface. These roots are perennial and show annual growth rings, and like the trunk or branches increase in diameter. This (along with soil erosion) is why many tree roots eventually become exposed or appear at the surface. Woody roots have lignin along with cellulose and hemicelluloses in their cell walls. Woody roots have an outer bark and contains suberin. Suberin gives bark a corky characteristic and "waterproofs" the tissues.
In drier soils, some tree species will form "striker roots" at intervals along the framework system. These roots grow vertically downward until they encounter an obstacle or soil with insufficient oxygen for growth. They will often branch and form a second layer of roots deeper in the soil. These roots function as water and food storage areas for the tree.
Another type of root is the adventitious root. These roots will often form spontaneously at the root collar from large woody roots. Although it is not known exactly what causes their formation, they usually develop as a result of injury or excessive soil and/or deep mulch piled against the trunk.
Root Functions
Tree roots and soils are a cooperative venture that generates healthy trees and healthy soils. Understanding how roots grow and utilize the soil is critical to proper tree management.
Root Forms
Roots survive and grow where adequate water is available, temperatures are warm, and oxygen is present. Roots are limited by oxygen content, anaerobic conditions, and water depth. Near the base of the tree, deep-growing roots can be found because they are oxygenated through fissures and cracks in the soil generated as a result of mechanical forces moving the crown and trunk swaying from the wind.
Roots go through several forms:
The taproot - When a seed germinates, the primary root, or radicle as it is called in the embryo, emerges from the seed coat. The taproot system develops from the primary root. It is comprised of one major root that is thicker at its junction with the trunk or stem and tapers toward the growing tip. Taproots penetrate deep into the soil and are often used for food storage. They form small lateral roots and are often deep and provide excellent anchorage for the tree. With increasing soil density downward, taproot growth is restricted and lateral root growth increases. On fibrous root systems, most taproots disappear after the seedling stage. Taproot systems are found in many dicots and gymnosperms. One research report has indicated that less than 5% of all trees have a tap root system.
Fibrous root systems consist of many lateral roots that grow horizontally to stabilize a tree. Many if not all fibrous root systems develop from small adventitious roots that develop from stem tissue soon after germination, particularly in monocots. Fibrous roots are shallow, wide spreading, numerous, and good for soil erosion control because they cling to soil particles and hold the soil in place.
A combination of both root systems occurs in most species that grow a taproot as a seedling until a certain distance or obstacle such as a rock is encountered, and then the root system changes and continues to grow as a fibrous system. This frequently occurs with nursery grown trees when the tap root from the seedling encounters the edge of the container or is air-pruned at the bottom of the container.
{kind=link}
Sloping Root Systems (also called heart root systems), have roots growing at many different angles from the horizontal to the vertical. Research shows that 15% of all trees have sloping root systems where the roots appear to radiate from a common point at the base of the trunk.
Aerial roots are roots that do not grow underground, but emerge from the trunk. There are a number of different types of aerial roots.
{kind=link}
- Prop roots, which generally arise at stem nodes, provide additional support for the tree. Prop roots are
found in corn and also in some aquatic tree species. When aerial roots grow toward the ground, they become prop roots. - Buttress roots are similar to prop roots, except they originate at the base of the stem, rather than further up the stem. They can be pretty impressive structures on some large tropical trees.
- Pneumatophores are negatively geotropic roots found on some trees in swampy areas; they grow above
the water line or tide line. These roots help absorb oxygen. They are common on mangroves. The
knees of bald cypress (Taxodium distichum) are similar to pneumatophores roots.
Propagation Roots – Many trees can produce "suckers" from roots. Suckers are adventitious woody shoots. Aspen groves, such as Pando in Utah's Fishlake National Forest, in which many trees are formed from one original tree, can cover large areas and all the trees in the grove grow from root suckers. All these trees are genetically identical.
Root Nodules – Some legumes and a few other plants produce root nodules, which contain bacteria or cyanobacteria that can "fix" nitrogen (make nitrogen into the nitrate form that is useful as a plant nutrient). When the bacteria enter the root, they induce the root to form "tumors" which are the nodules. The bacteria then grow and produce nitrogen forms in the nodules.
Root Systems
There are two basic types of roots, woody and non-woody.
Non-woody roots are found mostly in the upper few inches of soil. The primary function of these roots is to absorb water and nutrients. These are often called feeder roots. These roots also produce certain hormones (gibberelins and cytokinins) in the root meristem that are translocated through the tree to control growth and development. Non-woody roots have very little lignin in their cell walls. These roots live for a short time; from a few weeks to a year.
There are two types of structures that form on non-woody roots: root hairs and mycorrhizae.
1. Root hairs are organs that are extensions of epidermal cells. In some trees, particularly deciduous trees, the root hairs increase root surface area and increase nutrient and water uptake. Evergreen trees may not have root hairs but may possess mycorrhizae.
2. Mycorrhizae are organs composed of tree and fungus tissue, a composite structure. Mycorrhizae are a type of fungi that live on and in the feeder roots. These fungi do not cause any harm to the tree. In fact, for most species it is very beneficial for the tree to have this fungal association. Scientists have recently discovered that the tips of roots and mycorrhizae are involved in underground communication among trees and other plants.
Woody roots are large lateral roots that form at the flare or root collar (where the top of a root meets the bottom of the trunk). The primary purpose of woody roots are to support and anchor the tree. They also provide water and mineral transport as well as carbohydrate storage. These roots are distinct for each tree species and provide the framework for the tree's root system. The general direction for this framework is radial and horizontal and these roots are located 8 to 12 inches (20 – 30 cm) below the soil surface. These roots are perennial and show annual growth rings, and like the trunk or branches increase in diameter. This (along with soil erosion) is why many tree roots eventually become exposed or appear at the surface. Woody roots have lignin along with cellulose and hemicelluloses in their cell walls. Woody roots have an outer bark and contains suberin. Suberin gives bark a corky characteristic and "waterproofs" the tissues.
In drier soils, some tree species will form "striker roots" at intervals along the framework system. These roots grow vertically downward until they encounter an obstacle or soil with insufficient oxygen for growth. They will often branch and form a second layer of roots deeper in the soil. These roots function as water and food storage areas for the tree.
Another type of root is the adventitious root. These roots will often form spontaneously at the root collar from large woody roots. Although it is not known exactly what causes their formation, they usually develop as a result of injury or excessive soil and/or deep mulch piled against the trunk.
Root Functions
Tree roots and soils are a cooperative venture that generates healthy trees and healthy soils. Understanding how roots grow and utilize the soil is critical to proper tree management.
Root Physiology
Trees have two general systems that grow along an axis, the shoot system (with leaves and stems) above ground, and the root system below ground. Because root systems are unseen, many people do not think about them unless the roots have grown so much that sidewalks get cracked and lifted, or a drainage system needs cleaning because roots have invaded damaged drain pipes.
The roots of trees are required for anchorage, absorption of nutrients and water, and the storage of starch. The development of a root system is dependent upon the tree's genetics, the soil, and the environment. A tree's root system must balance its shoot system, but not by weight or dimensions. The root system must supply the shoot system with sufficient moisture and nutrients, and the shoots must manufacture enough food to support growth of the root system.
Trees require water, oxygen, carbon dioxide, light, nutrients, appropriate temperature, correct pH, physical space for growth processes, and an open soil surface area for replenishment of essential resources. Roots utilize space in the soil. The more space that is infiltrated by roots, the more potential resources (water and nutrients) are available, and this is directly related to tree health and growth.
Root Parts
Growth in trees represents an expansion of tissues into new spaces. For roots, the tips elongate and the tissues thicken in diameter. Roots develop internally rather than from buds as occurs on stems. To develop in this manner, a root has several parts. These parts are described as follows, from the youngest part of the root to the oldest part:
Root Cap – This part of the root is at its very tip. The cap protects the tip of the root as it is forced through the soil by the elongating tissue behind it. The root cap cells are coated with mucigel, a slime sheath that helps to lubricate the root as it penetrates through soil particles. These cells are constantly being replaced. A mature tree may have millions of root caps.
Apical Meristem – This part of the root generates the cells that form the root cap in front and the region of elongation behind this area. Researchers believe that the root cap and the apical meristem have the ability to sense an obstacle in the soil and change the root growth direction. The root can also change directions to seek moisture or nutrients. This is also the area where roots can connect with roots from other plants to transfer messages pertaining to disease and pest attacks.
Region of Elongation – Cells in their fixed positions elongate to reach mature dimensions in the region of elongation. The vacuole (a large sac of fluid within a cell) plays a major role in this process. It uses water pressure to push against the walls to stretch the cellulose fibers as the cells elongate. Virtually all increases in root length, occur in the elongation region. Active cell division forces the root through the soil against the mass of the tree. During this time of elongation, roots are also sensitive to gravity and respond with gravitropism.
Region of Root Hairs or Differentiation -- In this area, root cells develop a more mature form and are differentiated into the epidermis and cortex. Differentiation is when a cell changes its structure, such as when a parenchyma cell becomes enlarged to form a vessel; then it is no longer a parenchyma cell. The cell is said to be differentiated, and the process is the differentiation of cells. The outer layer of cells is the epidermis that contains a single layer of flattened cells with very little cuticle or suberin.
Root Hair – A root hair is the extension of a single epidermal cell. As the epidermal cells mature, the root hairs atrophy and are replaced by root hairs on newer cells in the early stages of maturation. Root hairs absorb water and elements dissolved in the water. Root hairs are organs that grow within days when water, temperature, and soluble essential elements are at optimum levels. Root hairs die and are shed after a few weeks. As root hairs and mycorrhizae atrophy, they add organic material to the soil. Non-woody roots shed dying and dead root hairs and epidermal cells. Soil microorganisms digest the shed cells and recycle elements essential for life. Root hairs do not become lateral roots. Root hairs and mycorrhizae are alive and growing in midwinter in the non-frozen soil below the frozen soils at the soil surface. Firs, redwoods, and Scots pine do not have root hairs. Instead, they absorb water and nutrients through the thin-walled epidermis. In contrast, some trees such as the redbud (Cercis) and honeylocust (Gleditsia) have root hairs that last for several years.
Rhizosphere – The soil area around the root tip and along the absorbing root-soil interface is called the rhizosphere. It is a zone about one millimeter thick surrounding the living root hairs and their mycorrhizae and hyphae. Constantly changing mixes of organisms inhabit the rhizosphere and surrounding soil. Bacteria, soil viruses, actinomycetes, fungi, protozoans, slime molds, algae, nematodes, enchytraeid worms, earthworms, millipedes, centipedes, insects, mites, snails, and small animals are in constant competition for the water, food, and space in the rhizosphere.
Root Cortex – This is composed of loosely packed round tissue cells with large diameters. Absorbed water moves readily between cells through the porous cell walls of the cortex parenchyma. Cortex cells typically contain amyloplasts, plastids (storage sacs) filled with starch granules that occur in seeds, roots, and stems. The pathway of interconnected cell walls facilitating water movement is generally regarded as the apoplast. An alternative water pathway, which carries water from cell to cell through plasmodesmata passing through the interiors of the cells, is called the symplast.
Endodermis – The inner layer of cortex is the endodermis. As roots mature, the endodermis becomes a barrier to further movement of water and minerals between cells.
Within the cortex are the xylem, phloem, and pericycle:
Symplast – Symplast is the network of highly ordered, connected, and living axial and radial parenchyma cells in sapwood and inner bark. The symplast stores energy reserves. The living protoplasm is contained in thin-walled cells called the parenchyma, which have small cell wall openings that act as tunnels where the protoplasm of one cell connects with the protoplasm of adjoining cells. The symplast stores energy reserves. The apoplast (dead fibers and tissues) stores bound water. As the symplast decreases, so does storage space, and as the storage of energy reserves decreases, so does the defense potential. Pathogens seem to know this very well. The symplast is made up of radial and axial parenchyma cells:
Cambium – In older parts of the root, another meristem forms between the xylem and phloem. This cambial zone is sometimes called the vascular cambium. It is rarely made up of a single layer of cells. Mitosis in the cambium produces new secondary xylem to the inside and secondary phloem to the outside. The cambium zone in roots is like an accordion; during the resting period it is closed and during the growing season it is open.
Secondary Root Growth
A cambium cylinder develops from parenchyma cells between xylem and phloem in the primary root stele. Once formed, the cambium produces xylem inward and phloem outward. Additional parenchyma cells form rays. Outer bark or periderm is mostly dead cells lined with a fatty substance called suberin or cork. The phellogen (bark cambium) is the outermost part of the symplast and the end of the phloem rays. The annual growth of "wood" and cork-like bark in secondary roots is very similar to stem secondary growth.
Lateral Roots
In the area behind the region of root hairs, lateral roots are formed by sending out a root cap, apical meristem tissue, etc. into a new area of soil. The ability of primary root tips to enter soil pores, further open soil pores, and elongate through soil pores is dependent upon the force generated by the root and the soil's resistance to penetration. Cell division and subsequent osmotic enlargement of each new cell generate root growth forces. Adequate water is required, as well as oxygen for respiration, because tree roots can consume large amounts of oxygen during elongation.
Root Functions
Woody supportive roots account for a small proportion of the tree's total root length and metabolic carbon demand. The larger woody supportive roots extending from the base of a tree are long-lived (often as old as the tree) and comprise most of the lateral root biomass.
In contrast, fine roots comprise only 5-10% of total root biomass, yet can account for up to 90% of the tree's total root length and metabolic activity. In many forests, the annual carbon cost for fine root system production may represent one of the largest carbon sinks in forested ecosystems.
Trees have two general systems that grow along an axis, the shoot system (with leaves and stems) above ground, and the root system below ground. Because root systems are unseen, many people do not think about them unless the roots have grown so much that sidewalks get cracked and lifted, or a drainage system needs cleaning because roots have invaded damaged drain pipes.
The roots of trees are required for anchorage, absorption of nutrients and water, and the storage of starch. The development of a root system is dependent upon the tree's genetics, the soil, and the environment. A tree's root system must balance its shoot system, but not by weight or dimensions. The root system must supply the shoot system with sufficient moisture and nutrients, and the shoots must manufacture enough food to support growth of the root system.
Trees require water, oxygen, carbon dioxide, light, nutrients, appropriate temperature, correct pH, physical space for growth processes, and an open soil surface area for replenishment of essential resources. Roots utilize space in the soil. The more space that is infiltrated by roots, the more potential resources (water and nutrients) are available, and this is directly related to tree health and growth.
Root Parts
Growth in trees represents an expansion of tissues into new spaces. For roots, the tips elongate and the tissues thicken in diameter. Roots develop internally rather than from buds as occurs on stems. To develop in this manner, a root has several parts. These parts are described as follows, from the youngest part of the root to the oldest part:
Root Cap – This part of the root is at its very tip. The cap protects the tip of the root as it is forced through the soil by the elongating tissue behind it. The root cap cells are coated with mucigel, a slime sheath that helps to lubricate the root as it penetrates through soil particles. These cells are constantly being replaced. A mature tree may have millions of root caps.
Apical Meristem – This part of the root generates the cells that form the root cap in front and the region of elongation behind this area. Researchers believe that the root cap and the apical meristem have the ability to sense an obstacle in the soil and change the root growth direction. The root can also change directions to seek moisture or nutrients. This is also the area where roots can connect with roots from other plants to transfer messages pertaining to disease and pest attacks.
Region of Elongation – Cells in their fixed positions elongate to reach mature dimensions in the region of elongation. The vacuole (a large sac of fluid within a cell) plays a major role in this process. It uses water pressure to push against the walls to stretch the cellulose fibers as the cells elongate. Virtually all increases in root length, occur in the elongation region. Active cell division forces the root through the soil against the mass of the tree. During this time of elongation, roots are also sensitive to gravity and respond with gravitropism.
Region of Root Hairs or Differentiation -- In this area, root cells develop a more mature form and are differentiated into the epidermis and cortex. Differentiation is when a cell changes its structure, such as when a parenchyma cell becomes enlarged to form a vessel; then it is no longer a parenchyma cell. The cell is said to be differentiated, and the process is the differentiation of cells. The outer layer of cells is the epidermis that contains a single layer of flattened cells with very little cuticle or suberin.
Root Hair – A root hair is the extension of a single epidermal cell. As the epidermal cells mature, the root hairs atrophy and are replaced by root hairs on newer cells in the early stages of maturation. Root hairs absorb water and elements dissolved in the water. Root hairs are organs that grow within days when water, temperature, and soluble essential elements are at optimum levels. Root hairs die and are shed after a few weeks. As root hairs and mycorrhizae atrophy, they add organic material to the soil. Non-woody roots shed dying and dead root hairs and epidermal cells. Soil microorganisms digest the shed cells and recycle elements essential for life. Root hairs do not become lateral roots. Root hairs and mycorrhizae are alive and growing in midwinter in the non-frozen soil below the frozen soils at the soil surface. Firs, redwoods, and Scots pine do not have root hairs. Instead, they absorb water and nutrients through the thin-walled epidermis. In contrast, some trees such as the redbud (Cercis) and honeylocust (Gleditsia) have root hairs that last for several years.
Rhizosphere – The soil area around the root tip and along the absorbing root-soil interface is called the rhizosphere. It is a zone about one millimeter thick surrounding the living root hairs and their mycorrhizae and hyphae. Constantly changing mixes of organisms inhabit the rhizosphere and surrounding soil. Bacteria, soil viruses, actinomycetes, fungi, protozoans, slime molds, algae, nematodes, enchytraeid worms, earthworms, millipedes, centipedes, insects, mites, snails, and small animals are in constant competition for the water, food, and space in the rhizosphere.
Root Cortex – This is composed of loosely packed round tissue cells with large diameters. Absorbed water moves readily between cells through the porous cell walls of the cortex parenchyma. Cortex cells typically contain amyloplasts, plastids (storage sacs) filled with starch granules that occur in seeds, roots, and stems. The pathway of interconnected cell walls facilitating water movement is generally regarded as the apoplast. An alternative water pathway, which carries water from cell to cell through plasmodesmata passing through the interiors of the cells, is called the symplast.
Endodermis – The inner layer of cortex is the endodermis. As roots mature, the endodermis becomes a barrier to further movement of water and minerals between cells.
Within the cortex are the xylem, phloem, and pericycle:
- Xylem – The xylem consists of large diameter vessels. Xylem is not wood; it is one of the transport tissues in vascular plants. It transports free water and substances dissolved in the water from absorbing non-woody roots to the leaves. When xylem is lignified it is then correctly called wood. Lignified means that high amounts of the natural "cement" called lignin is deposited within the cellulose strands in the cell walls. This makes the cell walls very tough. Having tough, lignified cell walls is a unique feature of trees.
- Phloem – Found in patches between the xylem arms, phloem is comprised of sieve elements and companion cells. Phloem is another transport tissue. It transports energy-containing substances (carbohydrates) made in leaves, to the roots and other growing parts of the tree.
- Pericycle – The pericycle consists of parenchyma cells just inside of the endodermis, forming the rest of the stele other than the xylem and phloem.
Symplast – Symplast is the network of highly ordered, connected, and living axial and radial parenchyma cells in sapwood and inner bark. The symplast stores energy reserves. The living protoplasm is contained in thin-walled cells called the parenchyma, which have small cell wall openings that act as tunnels where the protoplasm of one cell connects with the protoplasm of adjoining cells. The symplast stores energy reserves. The apoplast (dead fibers and tissues) stores bound water. As the symplast decreases, so does storage space, and as the storage of energy reserves decreases, so does the defense potential. Pathogens seem to know this very well. The symplast is made up of radial and axial parenchyma cells:
- radial parenchyma cells run perpendicular in the trunk.
- axial parenchyma cells run parallel to the trunk.
Cambium – In older parts of the root, another meristem forms between the xylem and phloem. This cambial zone is sometimes called the vascular cambium. It is rarely made up of a single layer of cells. Mitosis in the cambium produces new secondary xylem to the inside and secondary phloem to the outside. The cambium zone in roots is like an accordion; during the resting period it is closed and during the growing season it is open.
Secondary Root Growth
A cambium cylinder develops from parenchyma cells between xylem and phloem in the primary root stele. Once formed, the cambium produces xylem inward and phloem outward. Additional parenchyma cells form rays. Outer bark or periderm is mostly dead cells lined with a fatty substance called suberin or cork. The phellogen (bark cambium) is the outermost part of the symplast and the end of the phloem rays. The annual growth of "wood" and cork-like bark in secondary roots is very similar to stem secondary growth.
Lateral Roots
In the area behind the region of root hairs, lateral roots are formed by sending out a root cap, apical meristem tissue, etc. into a new area of soil. The ability of primary root tips to enter soil pores, further open soil pores, and elongate through soil pores is dependent upon the force generated by the root and the soil's resistance to penetration. Cell division and subsequent osmotic enlargement of each new cell generate root growth forces. Adequate water is required, as well as oxygen for respiration, because tree roots can consume large amounts of oxygen during elongation.
Root Functions
Woody supportive roots account for a small proportion of the tree's total root length and metabolic carbon demand. The larger woody supportive roots extending from the base of a tree are long-lived (often as old as the tree) and comprise most of the lateral root biomass.
In contrast, fine roots comprise only 5-10% of total root biomass, yet can account for up to 90% of the tree's total root length and metabolic activity. In many forests, the annual carbon cost for fine root system production may represent one of the largest carbon sinks in forested ecosystems.
General Root Growth
Root growth, density, and mass, vary with soil conditions. Roots utilize soil pore spaces for access to water and essential nutrients. Growing roots follow pathways of interconnected soil pores. Pores can be the space between soil particles, between structural blocks, plates, grains, soil prisms, along fracture lines, and the paths of decayed roots, worm and other animal diggings, etc.
Most root systems of trees grow in the top three feet of the soil. This is where oxygen, water, and organic matter are most prevalent. The top third of the root system feeds the tree and lower roots are mostly involved with water acquisition. Roots rarely grow below four feet although there are cases of some roots accessing deep water sources. Roots have been measured at depths of almost 175 feet (50 m) in some desert areas. However, the major portion of a tree's root system is in the top three feet of soil. This makes it easier to understand why trees can be easily uplifted during windstorms or other soil disturbances.
Roots need a minimum of 3% oxygen in the soil to stay alive. Twelve percent is needed for new roots to form. Fortunately, most healthy soils contain about 20% oxygen. However, if surface changes have occurred or if the soil became compacted or saturated and contains only 5% oxygen, the existing roots would survive but new roots could not grow and the tree would become stressed. Soil types can also influence root development. While clay soils might have insufficient oxygen levels at three feet deep, sandy soils could have 15% oxygen at five feet deep.
Studies have shown root spread to be 4 to 7 times the drip line distance (radius) of the tree. Other studies have shown that once a tree is established, many roots will have grown an eventual distance equal to approximately 3 times the tree's height. These are important facts to remember when applying herbicides, fertilizers, insecticides, heavy metals, hydrocarbons, biological toxins and other soil, weed or pest treatments around trees that will impact plant growth or are at levels exceeding the EPA's standards for soil contaminants.
The color of healthy, vigorous tree root tips is white to light yellow, with smooth surfaces. Root tips affected by poor aeration as in waterlogged soils are dark brown, purple or even black and have rough-looking surfaces.
Root – Shoot Balance
As mentioned previously, for most plants there is a balance between the root area and shoot area. Shoots must have adequate leaf surface area for photosynthesis for the entire tree and roots must have adequate surface area to provide water and nutrients for the tree and leaves. If roots are damaged, then shoot growth can be affected. Similarly, poor shoot growth limits the available fuel needed for root growth. Fortunately, plants are able to replace damaged roots with new root growth pretty rapidly, just as new leaves and shoots are readily produced after a storm.
The latest research indicates that an area of excitable cells is in the root tip or in the region of elongation, just behind the root tip which acts like a tiny brain. These sensors in the root tip enable the growing root to change direction to go around an obstacle or to find water and nutrients. In some cases, the root tip will alter course before it hits an underground obstacle, showing that trees are capable of “sensing or seeing” an obstacle in the soil. Researchers have predicted there may be something like a “brain” in root tips or region of elongation that has yet to be discovered. Research by this author has revealed that the removal of the root tip results in the cessation of all growth from that root. The root with the tip removed will not send out new roots to replace the missing root tip, but other roots will grow nearby to compensate for the missing or non-functioning root.
One root tip may not be able to do much, but most trees have millions of individual roots, each with a single root tip. Instead of a single large brain like people, trees may have millions of small brains or sensors that work together in a complex, leaderless network. The proof of this evolutionary observation is that a tree can survive and continue growing even after losing 95% or more of its roots during transplanting.
Water Uptake
Water enters the root through the root hairs. These extensions of epidermal cells have sticky walls and adhere tightly to soil particles with their film of moisture. Once within the epidermis, water passes through the cortex, mainly traveling between the cells. However, in order to enter the stele, it must pass through the cytoplasm of the cells of the endodermis. Once within the stele, water is free again to move between cells as well as through them. In young roots, water enters directly into the xylem. In older roots, it may have to pass first through a band of phloem and cambium. It does so by traveling through horizontally elongated cells, called the xylem rays. Water eventually makes its way to the leaf where it participates in the photosynthesis process, then leaves the tree though stomatal pores as water vapor. The rate of water loss is determined by temperature, wind, sunlight, and humidity. The hotter, drier, and windier a site, the faster the water vapor leaves the tree.
Mineral Uptake
Most researchers have indicated that minerals can move only if they are soluble. Water and essential elements from the soil move upward in the xylem-vessels known as tracheids. But other researchers have indicated that minerals enter separately. Even when no water is being absorbed, minerals enter freely. Minerals can enter against their concentration gradient; that is, by active transport. Anything that interferes with the metabolism of root cells interferes with mineral absorption. Plants absorb their nutrients in an inorganic form.
Organic fertilizers meet no nutritional need of the tree until their constituents have been degraded to inorganic forms. Organic matter does play an important role in making good soil texture, but only to the extent that it can yield inorganic ions; then it can meet the nutritional needs of the tree. Once within the epidermis, inorganic ions pass inward from cell to cell, probably through plasmodesmata. (Plasmodesmata is a narrow thread of cytoplasm that passes through the cell walls of adjacent plant cells and allows communication between them.) The final step from the cytoplasm of the pericycle cells to the xylem is probably accomplished once again by active transport.
Gas Exchange
The older parts of roots are sheathed in layers of dead cork cells impregnated with a waxy, waterproof and air proof substance called suberin. This sheath reduces water loss and is as impervious to oxygen and carbon dioxide as it is to water. However, the cork is perforated by nonsuberized pores called lenticels. These permit the exchange of oxygen and carbon dioxide between the air and the living cells beneath.
Compaction
Soil compaction is one of the biggest problems a tree root can have. Water and oxygen become unavailable when the pore spaces are closed. To prevent compaction, avoid large grade changes during construction, both cutting and filling. Also avoid the use of plastic as mulch or under mulches. Recent research has discovered that the pores of weed barrier fabrics can easily become clogged with molds and other microscopic residue also making the barriers unacceptable for soil health. A better solution is to increase the layer of mulch a couple of inches (5 cm) every couple of years.
For effective root growth, pore sizes in the soil must be larger than root tips. With compaction in a root colonization area, pore space diameters become smaller. Once soil pore diameters are less than the diameter of main root tips, many growth problems can occur. First, the main axis of a root becomes thicker to exert more force to squeeze into diminished sized pores. As roots thicken, growth slows and more laterals of various diameters are generated. Lateral root tip diameters begin initiation by a growth regulator and the extent of vascular tissue connections. If laterals are small enough to fit into the pore sizes of the compacted soil, then lateral growth will continue while the main axis of the root is constrained. If the soil pore sizes are too small for even the lateral roots, root growth will cease.
Tree Species Tolerance
Across the gene combinations which comprise tree forms, there is a great variability in reactions to soil compaction. A tree's ability to tolerate compacted soil conditions is associated with four primary internal mechanisms:
1. effective and fast reaction to mechanical damage,
2. continuation of respiration under chronic oxygen shortages,
3. ability to continue to turnover, reorient, and adjust absorbing root systems,
4. ability to deal with chemical toxins.
As there are many different soils and responses to compaction, so too are there many gradations of tree responses to compaction.
Root growth, density, and mass, vary with soil conditions. Roots utilize soil pore spaces for access to water and essential nutrients. Growing roots follow pathways of interconnected soil pores. Pores can be the space between soil particles, between structural blocks, plates, grains, soil prisms, along fracture lines, and the paths of decayed roots, worm and other animal diggings, etc.
Most root systems of trees grow in the top three feet of the soil. This is where oxygen, water, and organic matter are most prevalent. The top third of the root system feeds the tree and lower roots are mostly involved with water acquisition. Roots rarely grow below four feet although there are cases of some roots accessing deep water sources. Roots have been measured at depths of almost 175 feet (50 m) in some desert areas. However, the major portion of a tree's root system is in the top three feet of soil. This makes it easier to understand why trees can be easily uplifted during windstorms or other soil disturbances.
Roots need a minimum of 3% oxygen in the soil to stay alive. Twelve percent is needed for new roots to form. Fortunately, most healthy soils contain about 20% oxygen. However, if surface changes have occurred or if the soil became compacted or saturated and contains only 5% oxygen, the existing roots would survive but new roots could not grow and the tree would become stressed. Soil types can also influence root development. While clay soils might have insufficient oxygen levels at three feet deep, sandy soils could have 15% oxygen at five feet deep.
Studies have shown root spread to be 4 to 7 times the drip line distance (radius) of the tree. Other studies have shown that once a tree is established, many roots will have grown an eventual distance equal to approximately 3 times the tree's height. These are important facts to remember when applying herbicides, fertilizers, insecticides, heavy metals, hydrocarbons, biological toxins and other soil, weed or pest treatments around trees that will impact plant growth or are at levels exceeding the EPA's standards for soil contaminants.
The color of healthy, vigorous tree root tips is white to light yellow, with smooth surfaces. Root tips affected by poor aeration as in waterlogged soils are dark brown, purple or even black and have rough-looking surfaces.
Root – Shoot Balance
As mentioned previously, for most plants there is a balance between the root area and shoot area. Shoots must have adequate leaf surface area for photosynthesis for the entire tree and roots must have adequate surface area to provide water and nutrients for the tree and leaves. If roots are damaged, then shoot growth can be affected. Similarly, poor shoot growth limits the available fuel needed for root growth. Fortunately, plants are able to replace damaged roots with new root growth pretty rapidly, just as new leaves and shoots are readily produced after a storm.
The latest research indicates that an area of excitable cells is in the root tip or in the region of elongation, just behind the root tip which acts like a tiny brain. These sensors in the root tip enable the growing root to change direction to go around an obstacle or to find water and nutrients. In some cases, the root tip will alter course before it hits an underground obstacle, showing that trees are capable of “sensing or seeing” an obstacle in the soil. Researchers have predicted there may be something like a “brain” in root tips or region of elongation that has yet to be discovered. Research by this author has revealed that the removal of the root tip results in the cessation of all growth from that root. The root with the tip removed will not send out new roots to replace the missing root tip, but other roots will grow nearby to compensate for the missing or non-functioning root.
One root tip may not be able to do much, but most trees have millions of individual roots, each with a single root tip. Instead of a single large brain like people, trees may have millions of small brains or sensors that work together in a complex, leaderless network. The proof of this evolutionary observation is that a tree can survive and continue growing even after losing 95% or more of its roots during transplanting.
Water Uptake
Water enters the root through the root hairs. These extensions of epidermal cells have sticky walls and adhere tightly to soil particles with their film of moisture. Once within the epidermis, water passes through the cortex, mainly traveling between the cells. However, in order to enter the stele, it must pass through the cytoplasm of the cells of the endodermis. Once within the stele, water is free again to move between cells as well as through them. In young roots, water enters directly into the xylem. In older roots, it may have to pass first through a band of phloem and cambium. It does so by traveling through horizontally elongated cells, called the xylem rays. Water eventually makes its way to the leaf where it participates in the photosynthesis process, then leaves the tree though stomatal pores as water vapor. The rate of water loss is determined by temperature, wind, sunlight, and humidity. The hotter, drier, and windier a site, the faster the water vapor leaves the tree.
Mineral Uptake
Most researchers have indicated that minerals can move only if they are soluble. Water and essential elements from the soil move upward in the xylem-vessels known as tracheids. But other researchers have indicated that minerals enter separately. Even when no water is being absorbed, minerals enter freely. Minerals can enter against their concentration gradient; that is, by active transport. Anything that interferes with the metabolism of root cells interferes with mineral absorption. Plants absorb their nutrients in an inorganic form.
Organic fertilizers meet no nutritional need of the tree until their constituents have been degraded to inorganic forms. Organic matter does play an important role in making good soil texture, but only to the extent that it can yield inorganic ions; then it can meet the nutritional needs of the tree. Once within the epidermis, inorganic ions pass inward from cell to cell, probably through plasmodesmata. (Plasmodesmata is a narrow thread of cytoplasm that passes through the cell walls of adjacent plant cells and allows communication between them.) The final step from the cytoplasm of the pericycle cells to the xylem is probably accomplished once again by active transport.
Gas Exchange
The older parts of roots are sheathed in layers of dead cork cells impregnated with a waxy, waterproof and air proof substance called suberin. This sheath reduces water loss and is as impervious to oxygen and carbon dioxide as it is to water. However, the cork is perforated by nonsuberized pores called lenticels. These permit the exchange of oxygen and carbon dioxide between the air and the living cells beneath.
Compaction
Soil compaction is one of the biggest problems a tree root can have. Water and oxygen become unavailable when the pore spaces are closed. To prevent compaction, avoid large grade changes during construction, both cutting and filling. Also avoid the use of plastic as mulch or under mulches. Recent research has discovered that the pores of weed barrier fabrics can easily become clogged with molds and other microscopic residue also making the barriers unacceptable for soil health. A better solution is to increase the layer of mulch a couple of inches (5 cm) every couple of years.
For effective root growth, pore sizes in the soil must be larger than root tips. With compaction in a root colonization area, pore space diameters become smaller. Once soil pore diameters are less than the diameter of main root tips, many growth problems can occur. First, the main axis of a root becomes thicker to exert more force to squeeze into diminished sized pores. As roots thicken, growth slows and more laterals of various diameters are generated. Lateral root tip diameters begin initiation by a growth regulator and the extent of vascular tissue connections. If laterals are small enough to fit into the pore sizes of the compacted soil, then lateral growth will continue while the main axis of the root is constrained. If the soil pore sizes are too small for even the lateral roots, root growth will cease.
Tree Species Tolerance
Across the gene combinations which comprise tree forms, there is a great variability in reactions to soil compaction. A tree's ability to tolerate compacted soil conditions is associated with four primary internal mechanisms:
1. effective and fast reaction to mechanical damage,
2. continuation of respiration under chronic oxygen shortages,
3. ability to continue to turnover, reorient, and adjust absorbing root systems,
4. ability to deal with chemical toxins.
As there are many different soils and responses to compaction, so too are there many gradations of tree responses to compaction.
Soil Microorganisms
Trees grow their best in forest soils, but when trees are brought into the cities and planted in urban soil, they will not do well. The major cause of this among many others, is the lack of microorganisms in the soil. Soil microorganisms consist of animals such as algae, viruses, insects, earthworms, protozoa, arthropods, bacteria, fungi, and nematodes. They carry out numerous biological functions such as organic decomposition, create humus, suppress pathogens, improve soil properties and make nutrients available for plant growth.
Algae
Soil algae are filamentous or single-celled photo-autotrophic microorganisms. They use sunlight as an energy source and therefore their numbers are usually greatest near the surface. Soil algae assist in dissolving minerals and creating soil. Soil algae are everywhere and grow best under moist to wet conditions, though some are found in hot or cold deserts. Algae plus fungi form lichens, which are the important primary colonizers of rock. They are also components of microbial crusts in deserts.
Algae are grouped according to four common characteristics:
1. Diatoms have a siliceous cell wall that persists after death.
2. The eukaryotes group consists of a nucleus inside a cell membrane.
3. Prokaryotes are a group, formerly called blue-green algae that lack a cell nucleus.
4. The photo-autotrophs group can photosynthesize.
Bacteria
Bacteria decompose organic matter. Nitrogen gas from the air is chemically bound by bacteria into soluble or insoluble organic compounds that degrade in time, releasing soluble nitrogen compounds like ammonia. Bacterial nodules are common on many trees in the legume family. Rhizosphere bacteria are useful in legumes for fixing nitrogen to the roots. Nitrogen is fixed by bacteria in nodules. Most bacteria convert nitrogen into a form that is available to trees. Other bacteria denitrify nitrates while retaining nutrients like sulfur. Bacteria will also convert insoluble mineral phosphorus and iron into soluble products that trees can use.
Bacterial waste products become soil humus. Some bacteria are very useful in compost piles, while others will thrive in anaerobic composts, which generates odor and are harmful to plants. There are also some bacteria that cause diseases in trees. Bacteria often live in colonies of thousands of individuals and produce substances that hold soil particles together. A teaspoon of forest soil can contain 20 million to 2 billion bacteria.
Fungi
Fungi are the most important microorganisms in the soil for tree growth. Fungi are organisms that obtain their energy from other organisms, have a vegetative body made up of microscopic cells that may exist as individuals, but usually are joined to form long tube-like filaments called a mycelium. They reproduce by asexual or sexual spores and have well-defined nuclei. Fungi do not manufacture their own food nor do they have chlorophyll. Fungi include the yeast, molds, smuts, rusts, mushrooms, and many other forms that do not have common names. There are over 4,000 genera and 50,000 species known. They may dominate the biomass and metabolic activity in many soils. A teaspoon of forest soil can contain up to 40 miles of fungal hyphae. Fungi often depend upon living or dead organic matter for carbon and energy. Fungi are aerobic organisms but many can tolerate very low oxygen levels. Although many fungi are responsible for causing diseases such as rots and wilts, they are more important because they form a symbiotic relationship with a tree's roots. This association is called mycorrhizae. A tree cannot survive without mycorrhizae. Mycorrhizae are discussed in more detail below.
Fungi are the most versatile and persistent of any group who's job is decomposing microorganisms. They decompose cellulose, starch, gums, and lignin. They are more efficient in the assimilation of organic matter into their tissues (50%) than bacteria (20%). They continue to decompose resistant residues after bacteria and actinomycetes have ceased to function. Fungi are very important to soil fertility. They conduct nutrient cycling and improve soil tilth by stabilizing soil aggregates. They may produce mycotoxins, which are highly toxic to plants or animals and they may be predatory on other microorganisms. Fungi are especially important decomposers in acidic forest soils.
Tree roots function to absorb water and minerals however, the majority of vascular plants, including trees, form root associations with fungi to increase their absorption of mineral nutrients. Fungi, which live by absorbing nutrients from their surroundings, are ideal organisms to make these associations. The fungus obtains carbohydrates from the host tree, and in exchange, absorbs water and nutrients from the soil for the tree to use.
The life cycle of a fungus begins as a spore that grows when conditions are right. Out of the spore wall grows a hyphae that looks like a clear, microscopic fingertip. The body of the fungus is made up of a network of hyphal threads collectively called the mycelium. The mycelium grows in soil or within dead wood or living organisms. When growing conditions are favorable, the mycelium develop fruiting bodies, appearing as what we recognize as molds, mushrooms, or other forms. These fruiting bodies will produce new spores.
Penetrating hyphae branch off runner hyphae and infect fleshy young roots usually within a short distance of the root-growing tip. Once inside the root, these hyphae pierce root cell walls and form arbuscules between the root cell wall and cell membrane. Arbuscules are organs for exchanging nutrients with root cells. In addition to arbuscules, penetrating hyphae also produce storage organs called vesicles in spaces between root cells. Runner hyphae follow roots as they grow in the soil surrounding the infected root and absorb water and nutrients more efficiently than root hairs. The absorbed water and nutrients subsequently move to the penetrating hyphae and then into the infected root.
Roots attract mycorrhizal hyphae by exuding organic acids that act as chemical signals. As root tips continue to grow out into the soil, the infected root segment begins to harden. This hardening causes an end to the mycorrhizae and ensures that fungal colonization of roots occurs only on fleshy portions of the root system that are most active in absorbing water and nutrients.
Fungi decompose dead plants, pine needles, bark, wood, and animal matter. These saprophytes act as recyclers of dead organic matter, obtaining food from this material. Hyphal tips release enzymes that eventually decompose and release organic materials into the surrounding environment. Saprophytic fungi appear on dead trees, logs, tree litter such as leaves, and even on dead insects and animals. Fungal waste products that cannot be broken down further become soil humus.
Bacteria and Fungi
Bacteria and fungi perform important tasks that improve soil fertility. These tasks are typically performed in the root zone of host plants, which provides the sugar to fuel the process. Other bacteria and fungi, including mycorrhizal fungi and decomposing fungi like Tricoderma, carry out phosphorus solubilization. Biodegradation and recycling of organic matter is carried out by various bacteria and decomposing fungi, and is assisted by other microorganisms such as protozoa, insects, and earthworms. Both bacteria and mycorrhizal fungi exude glue-like materials that produce particle aggregates and contribute to good soil structure. This reduces compaction and increases porosity, as well as the soil's capacity to hold air and water. In addition, various microbes produce natural surfactants that promote water infiltration through the soil.
Mold Fungi
Mold fungi are distinctly filamentous, microscopic or submicroscopic, and they play an extremely important role in soil organic matter breakdown. Molds grow vigorously in acidic, neutral, and alkaline soils and they may dominate the micro flora in acidic surface soils. Four common genera are: Penicillium, Mucor, Fusarium, and Aspergillis. The complexity of organic matter seems to determine which of these particular molds are prevalent. Molds are tiny, with more or less than 100,000 to 1 million mold fungi per gram of soil.
Actinomycetes
Actinomycetes are microorganisms that have some characteristics of fungi and bacteria. Soil actinomycetes are very tolerant of water stress. Actinomycetes often give that "good earth" aroma after a rain.
Some benefits for trees:
1) Actinomycetes convert nitrogen in air to a usable form (fixed nitrogen),
2) Actinomycetes hold water (cell coatings).
Other Microorganisms
Protozoa, the most numerous one-celled organisms, are useful in decomposing organic matter and bacteria in the soil. They convert nitrogen into tree-available forms. They are also useful for destroying undesirable bacteria, nematodes, and some fungi, and there may be 100 to 100,000 protozoa per teaspoon of soil. To improve protozoan numbers, bacterial biomass needs to be enhanced. Protozoa inoculants are compost and some commercially available protozoan cultures. Protozoa come in three major groups, the ciliates, flagellates, and the amoebae. The relative numbers of these groups correspond to the oxygen level in the soil and whether the soil is aerobic or anaerobic.
Nematodes are microscopic worms that are tiny and round and are the most numerous multicellular animal on earth. The largest is just barely visible to the naked eye and there may be 5 to 500 beneficial nematodes per teaspoon of forest soil. Some nematodes will eat disease-causing fungi and some eat harmful nematodes. They also decompose organic matter. A bacterial feeding nematode will consume about 100 bacteria per day, while fungal feeding nematodes consume 80 feet (24m) of hyphae per day.
Pesticide Use
Any time nematicides, insecticides, fumigants, or herbicides are used, the beneficial nematodes will be killed and the root eating nematodes take over. Healthy soil contains predator nematodes and colonies of bacteria and fungi that make it difficult for the harmful nematodes to find roots. Compost is the only source of inoculants for the beneficial nematodes.
Arthropods
Arthropods are organisms that range from mites to beetles, centipedes, and wood lice. Their role in soil depends on which creature they are. Those with sucking mouth parts attack trees by sucking tree juices or sap. Those with jaws attack fungi or fungal feeders such as protozoa and nematodes. Arthropods do not decompose plant material; they just make it smaller for bacteria and fungi to decompose. They do make air passages in soil and form channels for water, roots, and other soil animals. Generally, soils disturbed by plowing, disking, chiseling, etc. will not have significant arthropod populations for a year or more unless mulch is placed on the soil surface. The principal role of these creatures is to recycle nutrients and make them available for plants and smaller microorganisms.
Actinorhizae
Actinorhizae fix nitrogen, which means that nitrogen in the air is converted to a form that can be used by the tree. In other words, the actinorhizae fix atmospheric pure nitrogen to ammonia, which can be chemically altered by some bacteria to nitrate ions. Actinorhizae are common on trees in many genera such as Alnus, Elaeagnus, Casuarina, to name a few.
Soil Enzymes
Soil enzymes are a group of proteins or conjugated proteins produced by living organisms and functioning as biochemical catalysts. Soil enzymes play an important role in soil microbial ecology by catalyzing reactions in soils and serve as an indicator of soil and ecosystem management in soil biology.
Earthworms
These small creatures carry organic matter from the surface deep into the soil while their holes provide a means of water and oxygen to percolate deep into the soil layers. Earthworms will eat anything they can get in their mouths. They will digest fungi, bacteria, protozoa, nematodes, and small arthropods. They fertilize the soil, promote tree growth, and neutralize soil acidity.
Glomalin
Glomalin is a unique fungal protein that holds soils together. The gooey protein is secreted through hyphae. Glomalin sloughs off of the hyphae and finds its way into soil. It coats soil particles and holds them together as aggregates or small grains of soil. Glomalin can be as high as 2% of the total weight of a soil aggregate.
Trees grow their best in forest soils, but when trees are brought into the cities and planted in urban soil, they will not do well. The major cause of this among many others, is the lack of microorganisms in the soil. Soil microorganisms consist of animals such as algae, viruses, insects, earthworms, protozoa, arthropods, bacteria, fungi, and nematodes. They carry out numerous biological functions such as organic decomposition, create humus, suppress pathogens, improve soil properties and make nutrients available for plant growth.
Algae
Soil algae are filamentous or single-celled photo-autotrophic microorganisms. They use sunlight as an energy source and therefore their numbers are usually greatest near the surface. Soil algae assist in dissolving minerals and creating soil. Soil algae are everywhere and grow best under moist to wet conditions, though some are found in hot or cold deserts. Algae plus fungi form lichens, which are the important primary colonizers of rock. They are also components of microbial crusts in deserts.
Algae are grouped according to four common characteristics:
1. Diatoms have a siliceous cell wall that persists after death.
2. The eukaryotes group consists of a nucleus inside a cell membrane.
3. Prokaryotes are a group, formerly called blue-green algae that lack a cell nucleus.
4. The photo-autotrophs group can photosynthesize.
Bacteria
Bacteria decompose organic matter. Nitrogen gas from the air is chemically bound by bacteria into soluble or insoluble organic compounds that degrade in time, releasing soluble nitrogen compounds like ammonia. Bacterial nodules are common on many trees in the legume family. Rhizosphere bacteria are useful in legumes for fixing nitrogen to the roots. Nitrogen is fixed by bacteria in nodules. Most bacteria convert nitrogen into a form that is available to trees. Other bacteria denitrify nitrates while retaining nutrients like sulfur. Bacteria will also convert insoluble mineral phosphorus and iron into soluble products that trees can use.
Bacterial waste products become soil humus. Some bacteria are very useful in compost piles, while others will thrive in anaerobic composts, which generates odor and are harmful to plants. There are also some bacteria that cause diseases in trees. Bacteria often live in colonies of thousands of individuals and produce substances that hold soil particles together. A teaspoon of forest soil can contain 20 million to 2 billion bacteria.
Fungi
Fungi are the most important microorganisms in the soil for tree growth. Fungi are organisms that obtain their energy from other organisms, have a vegetative body made up of microscopic cells that may exist as individuals, but usually are joined to form long tube-like filaments called a mycelium. They reproduce by asexual or sexual spores and have well-defined nuclei. Fungi do not manufacture their own food nor do they have chlorophyll. Fungi include the yeast, molds, smuts, rusts, mushrooms, and many other forms that do not have common names. There are over 4,000 genera and 50,000 species known. They may dominate the biomass and metabolic activity in many soils. A teaspoon of forest soil can contain up to 40 miles of fungal hyphae. Fungi often depend upon living or dead organic matter for carbon and energy. Fungi are aerobic organisms but many can tolerate very low oxygen levels. Although many fungi are responsible for causing diseases such as rots and wilts, they are more important because they form a symbiotic relationship with a tree's roots. This association is called mycorrhizae. A tree cannot survive without mycorrhizae. Mycorrhizae are discussed in more detail below.
Fungi are the most versatile and persistent of any group who's job is decomposing microorganisms. They decompose cellulose, starch, gums, and lignin. They are more efficient in the assimilation of organic matter into their tissues (50%) than bacteria (20%). They continue to decompose resistant residues after bacteria and actinomycetes have ceased to function. Fungi are very important to soil fertility. They conduct nutrient cycling and improve soil tilth by stabilizing soil aggregates. They may produce mycotoxins, which are highly toxic to plants or animals and they may be predatory on other microorganisms. Fungi are especially important decomposers in acidic forest soils.
Tree roots function to absorb water and minerals however, the majority of vascular plants, including trees, form root associations with fungi to increase their absorption of mineral nutrients. Fungi, which live by absorbing nutrients from their surroundings, are ideal organisms to make these associations. The fungus obtains carbohydrates from the host tree, and in exchange, absorbs water and nutrients from the soil for the tree to use.
The life cycle of a fungus begins as a spore that grows when conditions are right. Out of the spore wall grows a hyphae that looks like a clear, microscopic fingertip. The body of the fungus is made up of a network of hyphal threads collectively called the mycelium. The mycelium grows in soil or within dead wood or living organisms. When growing conditions are favorable, the mycelium develop fruiting bodies, appearing as what we recognize as molds, mushrooms, or other forms. These fruiting bodies will produce new spores.
Penetrating hyphae branch off runner hyphae and infect fleshy young roots usually within a short distance of the root-growing tip. Once inside the root, these hyphae pierce root cell walls and form arbuscules between the root cell wall and cell membrane. Arbuscules are organs for exchanging nutrients with root cells. In addition to arbuscules, penetrating hyphae also produce storage organs called vesicles in spaces between root cells. Runner hyphae follow roots as they grow in the soil surrounding the infected root and absorb water and nutrients more efficiently than root hairs. The absorbed water and nutrients subsequently move to the penetrating hyphae and then into the infected root.
Roots attract mycorrhizal hyphae by exuding organic acids that act as chemical signals. As root tips continue to grow out into the soil, the infected root segment begins to harden. This hardening causes an end to the mycorrhizae and ensures that fungal colonization of roots occurs only on fleshy portions of the root system that are most active in absorbing water and nutrients.
Fungi decompose dead plants, pine needles, bark, wood, and animal matter. These saprophytes act as recyclers of dead organic matter, obtaining food from this material. Hyphal tips release enzymes that eventually decompose and release organic materials into the surrounding environment. Saprophytic fungi appear on dead trees, logs, tree litter such as leaves, and even on dead insects and animals. Fungal waste products that cannot be broken down further become soil humus.
Bacteria and Fungi
Bacteria and fungi perform important tasks that improve soil fertility. These tasks are typically performed in the root zone of host plants, which provides the sugar to fuel the process. Other bacteria and fungi, including mycorrhizal fungi and decomposing fungi like Tricoderma, carry out phosphorus solubilization. Biodegradation and recycling of organic matter is carried out by various bacteria and decomposing fungi, and is assisted by other microorganisms such as protozoa, insects, and earthworms. Both bacteria and mycorrhizal fungi exude glue-like materials that produce particle aggregates and contribute to good soil structure. This reduces compaction and increases porosity, as well as the soil's capacity to hold air and water. In addition, various microbes produce natural surfactants that promote water infiltration through the soil.
Mold Fungi
Mold fungi are distinctly filamentous, microscopic or submicroscopic, and they play an extremely important role in soil organic matter breakdown. Molds grow vigorously in acidic, neutral, and alkaline soils and they may dominate the micro flora in acidic surface soils. Four common genera are: Penicillium, Mucor, Fusarium, and Aspergillis. The complexity of organic matter seems to determine which of these particular molds are prevalent. Molds are tiny, with more or less than 100,000 to 1 million mold fungi per gram of soil.
Actinomycetes
Actinomycetes are microorganisms that have some characteristics of fungi and bacteria. Soil actinomycetes are very tolerant of water stress. Actinomycetes often give that "good earth" aroma after a rain.
Some benefits for trees:
1) Actinomycetes convert nitrogen in air to a usable form (fixed nitrogen),
2) Actinomycetes hold water (cell coatings).
Other Microorganisms
Protozoa, the most numerous one-celled organisms, are useful in decomposing organic matter and bacteria in the soil. They convert nitrogen into tree-available forms. They are also useful for destroying undesirable bacteria, nematodes, and some fungi, and there may be 100 to 100,000 protozoa per teaspoon of soil. To improve protozoan numbers, bacterial biomass needs to be enhanced. Protozoa inoculants are compost and some commercially available protozoan cultures. Protozoa come in three major groups, the ciliates, flagellates, and the amoebae. The relative numbers of these groups correspond to the oxygen level in the soil and whether the soil is aerobic or anaerobic.
Nematodes are microscopic worms that are tiny and round and are the most numerous multicellular animal on earth. The largest is just barely visible to the naked eye and there may be 5 to 500 beneficial nematodes per teaspoon of forest soil. Some nematodes will eat disease-causing fungi and some eat harmful nematodes. They also decompose organic matter. A bacterial feeding nematode will consume about 100 bacteria per day, while fungal feeding nematodes consume 80 feet (24m) of hyphae per day.
Pesticide Use
Any time nematicides, insecticides, fumigants, or herbicides are used, the beneficial nematodes will be killed and the root eating nematodes take over. Healthy soil contains predator nematodes and colonies of bacteria and fungi that make it difficult for the harmful nematodes to find roots. Compost is the only source of inoculants for the beneficial nematodes.
Arthropods
Arthropods are organisms that range from mites to beetles, centipedes, and wood lice. Their role in soil depends on which creature they are. Those with sucking mouth parts attack trees by sucking tree juices or sap. Those with jaws attack fungi or fungal feeders such as protozoa and nematodes. Arthropods do not decompose plant material; they just make it smaller for bacteria and fungi to decompose. They do make air passages in soil and form channels for water, roots, and other soil animals. Generally, soils disturbed by plowing, disking, chiseling, etc. will not have significant arthropod populations for a year or more unless mulch is placed on the soil surface. The principal role of these creatures is to recycle nutrients and make them available for plants and smaller microorganisms.
Actinorhizae
Actinorhizae fix nitrogen, which means that nitrogen in the air is converted to a form that can be used by the tree. In other words, the actinorhizae fix atmospheric pure nitrogen to ammonia, which can be chemically altered by some bacteria to nitrate ions. Actinorhizae are common on trees in many genera such as Alnus, Elaeagnus, Casuarina, to name a few.
Soil Enzymes
Soil enzymes are a group of proteins or conjugated proteins produced by living organisms and functioning as biochemical catalysts. Soil enzymes play an important role in soil microbial ecology by catalyzing reactions in soils and serve as an indicator of soil and ecosystem management in soil biology.
Earthworms
These small creatures carry organic matter from the surface deep into the soil while their holes provide a means of water and oxygen to percolate deep into the soil layers. Earthworms will eat anything they can get in their mouths. They will digest fungi, bacteria, protozoa, nematodes, and small arthropods. They fertilize the soil, promote tree growth, and neutralize soil acidity.
Glomalin
Glomalin is a unique fungal protein that holds soils together. The gooey protein is secreted through hyphae. Glomalin sloughs off of the hyphae and finds its way into soil. It coats soil particles and holds them together as aggregates or small grains of soil. Glomalin can be as high as 2% of the total weight of a soil aggregate.
{kind=link}
Mycorrhizae
Mycorrhizal fungi are naturally occurring fungi that are found in abundance in undisturbed native soil environments in all the continents except Antarctica. Mycorrhizae (my-ka-RY-zee) are not exactly a root, and they are not exactly a fungus; they fall halfway between. The word in Greek means fungus “myco” and root “rhiza”. The specialized fungi that facilitate these relationships are collectively called mycorrhizal fungi, although they are often referred to as just mycorrhizae. They are important because they form a symbiotic relationship with about 95% of all terrestrial plants including virtually all trees. By definition, mycorrhizal fungi are highly specialized organisms that colonize the fine, non-woody roots of other plants and trees, acting as highly efficient extensions of the roots. By extending the root's functional area and efficiency, mycorrhizae can increase a tree's absorptive root surface many times over. This network of roots and fungal partners connect (or communicate according to recent research) with other trees to exchange vital nutrients and even information such as an impending insect attack.
There are about 8,000 mycorrhizal fungi species identified so far. They grow and reproduce themselves in ways far different than the world of plants. Structurally their cells can contain chitin, the same material found in the claws of mammals and shells of insects. In fact, mycorrhizal and other fungi are more closely related to animals than plants. Trees value the mycorrhizal relationship. However, if one mycorrhizal fungi should fail, the tree will easily adapt to another one.
Healthy soils contain vast repositories of mycorrhizal spores in the coarse organic matter near the soil surface, where they germinate under moist, aerated conditions. The mycorrhizal association occurs when the fungi grow in the rhizosphere, (the area around a root) and invade the outer layer of root cells. The fungi then act like root hairs to extract minerals and water from the soil. They also filter out heavy metals, which are less detrimental to the fungi than to the tree's roots. The diverted pollutants then end up as fruiting bodies or mushrooms. While inoculated roots tend to be shorter and more branched than uninfected roots, their associated mycorrhizae act like root hairs on steroids, extending far beyond the root mass to exploit soil resources. Even in poorer soils, the fungi send their filaments (called mycelium) into the surrounding soil, effectively increasing the tree's roots and absorbing capacity from ten to several thousand times. As they explore the soil in search of nutrients more efficiently than root hairs, they translocate nutrients and moisture back to the root for absorption and use by the tree. In exchange, the tree will pay for this service by providing the fungus with carbohydrates, vitamins, and amino acids from photosynthesis. Mycorrhizal filaments are much smaller than roots, so they can easily penetrate into smaller spaces between soil particles. Mycorrhizae will not perform efficiently in highly fertilized soils. Therefore the tree has to rely on less efficient root hairs, which reduce the effectiveness of the fertilizer. A soil full of mycorrhizal fungi seldom has bad fungi or disease issues.
The rhizosphere is a huge habitat in the soil, because plants have so many root fibers. At least 100 million organisms live in each gram of soil, but in the rhizosphere, there may be a trillion organisms per gram of soil.
Benefits of Mycorrhizae
Compared to non-colonized roots, mycorrhizal roots have been shown to:
Better phosphorus absorption often plays a significant role in the growth enhancement effects of roots infection by mycorrhizal fungi. Research has shown that the hyphae can deliver up to 80% of a host tree's phosphorus
requirements.
Hyphae
As already noted, hyphae are the filamentous structures of fungi. They resemble long, hollow tubes, but some are only 1/25th the diameter of a human hair, yet can grow as long as 18 to 24 inches (45-60 cm) long. The mycorrhizal hyphae growing from the roots of one tree can, and often do, connect with mycorrhizal hyphae originating from another tree, which may even be a different species. This creates an underground network (and communication links) through which soil resources can be shared. The hyphae of AMF arbuscular mycorrhizae are coated with a special high-carbon glycol-protein called glomalin. As mentioned above, glomalin is a very sticky, adhesive, and durable compound. As the hyphae from these fungi grow through the soil, they tend to grow around soil particles, wrapping and binding them together into tiny aggregates using glomalin as a glue. Over time, a rich,
well-textured topsoil is created through this process.
Types of Mycorrhizae
Mycorrhizae can be divided into two groups that represent the most common and abundant forms:
Ectomycorrhizae is a group of fungi found on short, lateral roots. Ecto means outside, so ectomycorrhizal fungi form a mantle or sheath around the outside of the cortical root cells forming a thick coat of hyphae around the secondary roots of trees, transporting water and minerals from the soil into the roots. The white, web-like hyphae of ectomycorrhizae are often visible to the naked eye. This relationship occurs in certain groups of trees: most conifers (pines,hemlocks, spruces) and some hardwoods (oaks, beeches, chestnuts, hickories, birches, pecans, hazelnuts, and a few others). Depending on the type of mycorrhizal relationship, infected trees have the advantage of being able to obtain water and nutrients from tiny pores and crevices of soil, that non-mycorrhizal trees cannot access. Ectomycorrhizal fungi species tend to be somewhat more host specific than arbuscular mycorrhizal fungi (AMF). Therefore, if a mycorrhizal inoculant is to be considered, some care should be taken to select an inoculum that contains the appropriate fungal species known to colonize well with the target trees. Even with the appropriate fungus, environmental factors such as fertilization, liming, pesticide use, and topsoil removal can adversely affect the beneficial effects of the fungus. Based on my experience when planting a tree, I will take a shovel full of soil from around the roots of a similar species tree nearby and sprinkle the soil over the roots of a new planting rather than purchasing an inoculum that did not produce any positive results. The majority of ectomycorrhizae grow best between 64 and 80 degrees F (18 ー 27 degrees C), though some have a wider temperature tolerance. Pisolithus tinctorius, for example, can grow at soil temperatures of 93 F (34 C) or higher and is a good choice for appropriate host trees in warm climates.
Arbuscular Mycorrhizae is commonly referred to as AM, VAM, or endomycorrhizae. Endo means inside, and this is where the hyphae enter inside the root. They are found in virtually all terrestrial habitats that can support plants, regardless of the soil type, climate, or other growing conditions. They will associate with the roots of approximately 80% of all known terrestrial plants, including the vast majority of cultivated species and are found in abundance on all continents except Antarctica. Arbuscular mycorrhizal fungi populations are often lost in urban and suburban environments due to excavation and other major soil disturbances during development and construction. This modification can take place in the soil environment, particularly under landscape irrigation, which reduces arbuscular fungi colonization on some tree roots. Again, based on my experience when planting a tree, I will take a shovel full of soil from around the roots of a similar species tree nearby and sprinkle the soil over the roots of a new planting rather than purchasing an inoculum that did not produce any positive results. Therefore inoculation at planting time, through irrigation or by soil injection are useful strategies to re-introduce these organisms to the roots of trees and other landscape plants. Arbuscular fungi can reduce transplant shock, increase nutrient uptake efficiency, improve early growth, improve drought tolerance, and suppress fungal root pathogens in colonized trees. Ultimately, the ability of arbuscular fungi to increase tree growth is likely a function of tree species and other factors such as water availability and enhanced phosphorus uptake. Arbuscular fungi are important components of terrestrial plant communities and have been found to have complex mutual relationships with many of the microorganisms found in the rhizosphere and surrounding soils.
Urban Soil
Urban soils are generally missing many key soil components, including mycorrhizae. This is a soil that has been dug up, stripped of topsoil, compacted, and generally abused for plant growth in order to make it stable for buildings, sidewalks, and roadway construction. These are the places where the introduction of mycorrhizal fungi are really needed if trees are expected to grow. Since the 1990's, numerous companies have manufactured mycorrhizal inoculants containing fungal propagules and spores in a variety of products. Many urban planting projects have been planted with these commercial mycorrhizal products serving as liquid root dip or soil injection. However, these products have not been proven to be reliable in an average tree planting operation. Put simply, the commercial inoculants are products that lack scientific trials and proof of effectiveness. There are thousands of scientific papers on mycorrhizae and commercial inoculants but most have been written by people with a vested interest in the product – such as scientists who work for the commercial manufacturer.
Independent research does not always indicate that trees receiving root inoculations show greater growth than the control trees. Furthermore, according to inoculant manufacturers, inoculation via the soil injection method is particularly effective for larger established trees in compacted soils. Of course, results are most noticeable in highly disturbed soils, where native mycorrhizae populations are low or non-existent. Inoculation with mycorrhizal fungi at planting time may help trees get established and may help them survive with less water and limited nutrient resources. An inoculum containing multiple mycorrhizal species will provide diversity and therefore generally better results than one containing only one or two species. Some tree planters, this author included, have been known to simply add a shovel full of local forest duff taken from near some existing roots, and sprinkle it over the roots of the trees in their planting holes. Forest soils contain plenty of native fungi in the ground. The soil should come from nearby sites where similar trees are doing well. A shovel-full of rich compost may also be more effective than the commercial inoculants.
It is very difficult to determine if an inoculant is of high quality. Some manufacturers count 'propagules' instead of 'spores.' Propagules can include root fragments and other inert materials, so the spore count might actually be much lower. Furthermore, these very specific species almost certainly are from an area that is foreign to the soil where a tree is to be installed. It is even possible that these foreign species will inhibit the growth of the native species but there is often no way of knowing the mycorrhizal species in the product because the information on the packaging can be vague or confusing. In addition, it is never a good idea to import foreign species of any kind into a planting site.
How do you know your trees will benefit from the 2 or 3 types of fungi you buy? There is no way to know that a product contains active fungi. They could all be dead. Manufactured mycorrhizal fungi inoculants are heat sensitive. If the container is left in a building or a transport truck too long, the sustained high temperatures kill the fungi.
Most plants form mycorrhizal fungi associations without any human interference. Even in disturbed soils, mycorrhizal fungi will be introduced naturally over time as wind and animals carry fungal spores into the area.
Introducing mycorrhizae into urban soils may help but the species introduced will likely be different than what the trees will eventually develop on their own. Therefore a wide range of mycorrhizae species should be introduced instead of just a couple of species with an inoculum.
Some other examples are discussed in Seminar #73.
Nursery Production
Until recently, nurserymen did not know much about mycorrhizal fungi and did not try to add them to the nursery and landscaped soils. The soil-less media found in most nursery trays and containers contain no mycorrhizal fungi or spores. Therefore plants grown in these materials are not colonized. Often the
soil into which they will be transplanted are devoid of fungi. Inoculation of plants during greenhouse propagation will infect the roots with mycorrhizae which then remain in place after transplanting. It is relatively easy and economical to treat plants in the greenhouse. Once colonized, the benefits imparted remain with the plant permanently for the life of the plant or tree.
Mycorrhizal fungi are naturally occurring fungi that are found in abundance in undisturbed native soil environments in all the continents except Antarctica. Mycorrhizae (my-ka-RY-zee) are not exactly a root, and they are not exactly a fungus; they fall halfway between. The word in Greek means fungus “myco” and root “rhiza”. The specialized fungi that facilitate these relationships are collectively called mycorrhizal fungi, although they are often referred to as just mycorrhizae. They are important because they form a symbiotic relationship with about 95% of all terrestrial plants including virtually all trees. By definition, mycorrhizal fungi are highly specialized organisms that colonize the fine, non-woody roots of other plants and trees, acting as highly efficient extensions of the roots. By extending the root's functional area and efficiency, mycorrhizae can increase a tree's absorptive root surface many times over. This network of roots and fungal partners connect (or communicate according to recent research) with other trees to exchange vital nutrients and even information such as an impending insect attack.
There are about 8,000 mycorrhizal fungi species identified so far. They grow and reproduce themselves in ways far different than the world of plants. Structurally their cells can contain chitin, the same material found in the claws of mammals and shells of insects. In fact, mycorrhizal and other fungi are more closely related to animals than plants. Trees value the mycorrhizal relationship. However, if one mycorrhizal fungi should fail, the tree will easily adapt to another one.
Healthy soils contain vast repositories of mycorrhizal spores in the coarse organic matter near the soil surface, where they germinate under moist, aerated conditions. The mycorrhizal association occurs when the fungi grow in the rhizosphere, (the area around a root) and invade the outer layer of root cells. The fungi then act like root hairs to extract minerals and water from the soil. They also filter out heavy metals, which are less detrimental to the fungi than to the tree's roots. The diverted pollutants then end up as fruiting bodies or mushrooms. While inoculated roots tend to be shorter and more branched than uninfected roots, their associated mycorrhizae act like root hairs on steroids, extending far beyond the root mass to exploit soil resources. Even in poorer soils, the fungi send their filaments (called mycelium) into the surrounding soil, effectively increasing the tree's roots and absorbing capacity from ten to several thousand times. As they explore the soil in search of nutrients more efficiently than root hairs, they translocate nutrients and moisture back to the root for absorption and use by the tree. In exchange, the tree will pay for this service by providing the fungus with carbohydrates, vitamins, and amino acids from photosynthesis. Mycorrhizal filaments are much smaller than roots, so they can easily penetrate into smaller spaces between soil particles. Mycorrhizae will not perform efficiently in highly fertilized soils. Therefore the tree has to rely on less efficient root hairs, which reduce the effectiveness of the fertilizer. A soil full of mycorrhizal fungi seldom has bad fungi or disease issues.
The rhizosphere is a huge habitat in the soil, because plants have so many root fibers. At least 100 million organisms live in each gram of soil, but in the rhizosphere, there may be a trillion organisms per gram of soil.
Benefits of Mycorrhizae
Compared to non-colonized roots, mycorrhizal roots have been shown to:
- grow faster,
- live longer,
- be more physiologically active and energy efficient,
- absorb water and accumulate nutrients better,
- improve uptake of essential nutrients such as phosphorus and micro-nutrients,
- make trees more resistant to drought,
- be more tolerant of soil compaction,
- be more tolerant of high temperatures,
- be more tolerant to salt and high salinity soils,
- be more tolerant of pH extremes, provide better support for flower, seed, and fruit production,
- resist transplant shock and improve plant survival and establishment,
- resist and suppress fungal root pathogens and parasitic nematodes,
- be more tolerant to organic and inorganic soil toxins,
- enhance the biological conversion of atmospheric nitrogen to plant available nitrogen,
- provide access and absorb “bound-up” soil nutrients, such as many phosphorous compounds.
Better phosphorus absorption often plays a significant role in the growth enhancement effects of roots infection by mycorrhizal fungi. Research has shown that the hyphae can deliver up to 80% of a host tree's phosphorus
requirements.
Hyphae
As already noted, hyphae are the filamentous structures of fungi. They resemble long, hollow tubes, but some are only 1/25th the diameter of a human hair, yet can grow as long as 18 to 24 inches (45-60 cm) long. The mycorrhizal hyphae growing from the roots of one tree can, and often do, connect with mycorrhizal hyphae originating from another tree, which may even be a different species. This creates an underground network (and communication links) through which soil resources can be shared. The hyphae of AMF arbuscular mycorrhizae are coated with a special high-carbon glycol-protein called glomalin. As mentioned above, glomalin is a very sticky, adhesive, and durable compound. As the hyphae from these fungi grow through the soil, they tend to grow around soil particles, wrapping and binding them together into tiny aggregates using glomalin as a glue. Over time, a rich,
well-textured topsoil is created through this process.
Types of Mycorrhizae
Mycorrhizae can be divided into two groups that represent the most common and abundant forms:
Ectomycorrhizae is a group of fungi found on short, lateral roots. Ecto means outside, so ectomycorrhizal fungi form a mantle or sheath around the outside of the cortical root cells forming a thick coat of hyphae around the secondary roots of trees, transporting water and minerals from the soil into the roots. The white, web-like hyphae of ectomycorrhizae are often visible to the naked eye. This relationship occurs in certain groups of trees: most conifers (pines,hemlocks, spruces) and some hardwoods (oaks, beeches, chestnuts, hickories, birches, pecans, hazelnuts, and a few others). Depending on the type of mycorrhizal relationship, infected trees have the advantage of being able to obtain water and nutrients from tiny pores and crevices of soil, that non-mycorrhizal trees cannot access. Ectomycorrhizal fungi species tend to be somewhat more host specific than arbuscular mycorrhizal fungi (AMF). Therefore, if a mycorrhizal inoculant is to be considered, some care should be taken to select an inoculum that contains the appropriate fungal species known to colonize well with the target trees. Even with the appropriate fungus, environmental factors such as fertilization, liming, pesticide use, and topsoil removal can adversely affect the beneficial effects of the fungus. Based on my experience when planting a tree, I will take a shovel full of soil from around the roots of a similar species tree nearby and sprinkle the soil over the roots of a new planting rather than purchasing an inoculum that did not produce any positive results. The majority of ectomycorrhizae grow best between 64 and 80 degrees F (18 ー 27 degrees C), though some have a wider temperature tolerance. Pisolithus tinctorius, for example, can grow at soil temperatures of 93 F (34 C) or higher and is a good choice for appropriate host trees in warm climates.
{kind=link}
Arbuscular Mycorrhizae is commonly referred to as AM, VAM, or endomycorrhizae. Endo means inside, and this is where the hyphae enter inside the root. They are found in virtually all terrestrial habitats that can support plants, regardless of the soil type, climate, or other growing conditions. They will associate with the roots of approximately 80% of all known terrestrial plants, including the vast majority of cultivated species and are found in abundance on all continents except Antarctica. Arbuscular mycorrhizal fungi populations are often lost in urban and suburban environments due to excavation and other major soil disturbances during development and construction. This modification can take place in the soil environment, particularly under landscape irrigation, which reduces arbuscular fungi colonization on some tree roots. Again, based on my experience when planting a tree, I will take a shovel full of soil from around the roots of a similar species tree nearby and sprinkle the soil over the roots of a new planting rather than purchasing an inoculum that did not produce any positive results. Therefore inoculation at planting time, through irrigation or by soil injection are useful strategies to re-introduce these organisms to the roots of trees and other landscape plants. Arbuscular fungi can reduce transplant shock, increase nutrient uptake efficiency, improve early growth, improve drought tolerance, and suppress fungal root pathogens in colonized trees. Ultimately, the ability of arbuscular fungi to increase tree growth is likely a function of tree species and other factors such as water availability and enhanced phosphorus uptake. Arbuscular fungi are important components of terrestrial plant communities and have been found to have complex mutual relationships with many of the microorganisms found in the rhizosphere and surrounding soils.
{kind=link}
Urban Soil
Urban soils are generally missing many key soil components, including mycorrhizae. This is a soil that has been dug up, stripped of topsoil, compacted, and generally abused for plant growth in order to make it stable for buildings, sidewalks, and roadway construction. These are the places where the introduction of mycorrhizal fungi are really needed if trees are expected to grow. Since the 1990's, numerous companies have manufactured mycorrhizal inoculants containing fungal propagules and spores in a variety of products. Many urban planting projects have been planted with these commercial mycorrhizal products serving as liquid root dip or soil injection. However, these products have not been proven to be reliable in an average tree planting operation. Put simply, the commercial inoculants are products that lack scientific trials and proof of effectiveness. There are thousands of scientific papers on mycorrhizae and commercial inoculants but most have been written by people with a vested interest in the product – such as scientists who work for the commercial manufacturer.
Independent research does not always indicate that trees receiving root inoculations show greater growth than the control trees. Furthermore, according to inoculant manufacturers, inoculation via the soil injection method is particularly effective for larger established trees in compacted soils. Of course, results are most noticeable in highly disturbed soils, where native mycorrhizae populations are low or non-existent. Inoculation with mycorrhizal fungi at planting time may help trees get established and may help them survive with less water and limited nutrient resources. An inoculum containing multiple mycorrhizal species will provide diversity and therefore generally better results than one containing only one or two species. Some tree planters, this author included, have been known to simply add a shovel full of local forest duff taken from near some existing roots, and sprinkle it over the roots of the trees in their planting holes. Forest soils contain plenty of native fungi in the ground. The soil should come from nearby sites where similar trees are doing well. A shovel-full of rich compost may also be more effective than the commercial inoculants.
It is very difficult to determine if an inoculant is of high quality. Some manufacturers count 'propagules' instead of 'spores.' Propagules can include root fragments and other inert materials, so the spore count might actually be much lower. Furthermore, these very specific species almost certainly are from an area that is foreign to the soil where a tree is to be installed. It is even possible that these foreign species will inhibit the growth of the native species but there is often no way of knowing the mycorrhizal species in the product because the information on the packaging can be vague or confusing. In addition, it is never a good idea to import foreign species of any kind into a planting site.
How do you know your trees will benefit from the 2 or 3 types of fungi you buy? There is no way to know that a product contains active fungi. They could all be dead. Manufactured mycorrhizal fungi inoculants are heat sensitive. If the container is left in a building or a transport truck too long, the sustained high temperatures kill the fungi.
Most plants form mycorrhizal fungi associations without any human interference. Even in disturbed soils, mycorrhizal fungi will be introduced naturally over time as wind and animals carry fungal spores into the area.
Introducing mycorrhizae into urban soils may help but the species introduced will likely be different than what the trees will eventually develop on their own. Therefore a wide range of mycorrhizae species should be introduced instead of just a couple of species with an inoculum.
Some other examples are discussed in Seminar #73.
Nursery Production
Until recently, nurserymen did not know much about mycorrhizal fungi and did not try to add them to the nursery and landscaped soils. The soil-less media found in most nursery trays and containers contain no mycorrhizal fungi or spores. Therefore plants grown in these materials are not colonized. Often the
soil into which they will be transplanted are devoid of fungi. Inoculation of plants during greenhouse propagation will infect the roots with mycorrhizae which then remain in place after transplanting. It is relatively easy and economical to treat plants in the greenhouse. Once colonized, the benefits imparted remain with the plant permanently for the life of the plant or tree.
Plant Growth Regulators
Plant hormones, or plant growth regulators, are chemicals produced by plants that alter growth patterns. They can be found in many cells and tissues, although plant hormones seem to be concentrated in meristems and buds. One should not confuse hormones with enzymes. Any chemical reaction that occurs in a cell requires a specific catalyst. Enzymes are the catalysts found in cells. Hormones inhibit as well as promote cellular activities by sending chemical signals to cells.
The hormones identified in plants most often regulate division, elongation, and differentiation of cells. Plant hormones work in very small concentrations, affecting membrane properties, controlling gene expression, and affecting enzyme activity. In most cases, the effect plant hormones have on the plant depends on the location and concentration of the hormone relative to other hormones. Hormones often work in conjunction with each other, have overlapping effects, and work with environmental stimuli. There are several classes of plant hormones, including a number of recently “discovered” ones.
Auxins
The concept of chemical messengers in plants was proposed by Charles Darwin in 1881, who spent time looking at the phenomenon of phototropism in wheat seedlings. When a seedling is illuminated from the side, the shoot will bend towards the light. Darwin did a number of experiments and determined that a certain chemical traveled to the region of elongation and effected a differential elongation of cells furthest from the light sources. The chemical was subsequently studied and named auxin. Chemically, auxin is indoleacetic acid (IAA). A primary site of auxin production is the apical shoot meristem.
The most studied function of auxin is cell elongation when protons stimulate proteins that disrupt hydrogen bonds and break cross linkages in cellulose. This facilitates wall expansion when cells take in more water.
Cytokinins
Cytokinins are one of the molecules in DNA. Biologists have yet to identify the genes for the cytokinins found in plants. One hypothesis today is that cytokinins are actually made by symbiotic methyl bacteria that live within tree tissues. They are found primarily in root meristems, embryos, and fruits, and migrate from roots to the shoot systems of plants in xylem tissue.
The effects of cytokinins are often studied in tissue culture. Parenchyma cells grown in tissue culture will not divide and differentiate unless cytokinins and auxin are present. Cytokinins can promote auxiliary bud growth by overriding the inhibiting effect of auxin. This is one of the ways in which plants balance root and shoot growth.
Cytokinins also retard leaf death, by stimulating RNA and protein synthesis and delaying degradation of chlorophyll. Galls in plants take advantage of cytokinin function. The bacteria that form these tumors contain genes for the synthesis of cytokinins, which promote rapid cell growth.
Gibberellins
The discovery of gibberellins found that a fungus was responsible for abnormal growth. The fungus was Gibberella fujikuroi. Over 100 different gibberellins are known and are produced in roots and younger leaves. Gibberellins work with auxins to promote rapid elongation and division of stem tissue. This is seen in:
PBZ (Paclobutrazol)
PBZ is a gibberelin biosynthesis inhibitor that reduces the growth of many species and is commonly used on trees under utility lines or anywhere else the tree size needs to be controlled. The product has also been used to stimulate root regeneration after transplanting. It has been studied for the purpose of stabilizing declining trees that have insufficient fine root development. Treatment should be part of a complete tree care program including mulching.
Abscissic Acid
Abscissic acid (ABA) is a hormone that functions by inhibiting growth activities in times of environmental stress rather than by promoting growth. It got its name from the erroneous belief that it promoted the formation of abscission layers in leaves and fruits. It does not do this, although leaf abscission accompanies dormancy in many plants.
ABA promotes seed dormancy activities. ABA levels are high when seeds mature, promoting lowered metabolism and synthesis of proteins needed to withstand the dehydration associated with dormancy. Seeds germinate when ABA is degraded by some environmental incident such as when rain has washed the ABA out of the seed coat or light-stimulated enzymes. Breaking dormancy is relative to the ratio of ABA (which keeps seeds in dormancy) and gibberellins (which promote germination). Low levels of ABA in maturing seeds promotes premature germination.
ABA promotes stomata closure during leaf water deficit conditions. ABA in this case originates in roots, and detects low water level in root tissues, then moves upward and activates stomatal closure.
ABA derivatives, called dormins, are used in commercial nurseries to keep materials to be shipped in dormant conditions. The dormancy can be reversed with gibberellins.
Ethylene
Ethylene is a gas best known as a fruit ripener. Gardeners in China knew centuries ago that fruits ripened faster in rooms with burning incense. Citrus growers used kerosene stoves in the rooms in which they ripened their fruit. Today, grocer warehouses have ethylene rooms for ripening most of our produce. Ethylene affects many aspects of growth and development in tissues throughout the tree, but emphasis is on fruit maturation, leaf abscission, and senescence.
A shoot tip that encounters an immovable object will grow around the object by changing its growth direction. This occurs through differential elongation of cell walls as ethylene is synthesized, slowing cell wall expansion.
Brassinolides (Brassinosteroids)
Brassinolides is a tree steroid discovered in pollen of members of the mustard family, and is best studied in the Arabadopsis plant. Chemically they are very similar to animal steroid hormones. Brassinosteroids signal cell elongation and cell division. Brassinosteroids promote differentiation of xylem tissue. Brassinosteroids can also retard leaf abscission. Absence of brassinolides results in dwarf plants.
Salicylic Acid
Salicylic acid is known to activate defense genes against pathogen invaders. Salicylic acid, a phenolic extract from Salix (willow) bark, was long used as an analgesic. It is now prepared commercially and is the active ingredient of aspirin.
Jasmonates
Jasmonates are a group of fatty acid derivatives. They appear to have a role in seed germination, root growth, and the storage of protein in seeds. Synthesis of defense proteins may be triggered by jasmonates.
Systemin
Systemin is a small peptide found in wound tissue. It may stimulate defense activities in other parts of the tree to prevent more wounding.
Oligosaccharins
Oligosaccharins are short chain sugars in cell walls that may have a role in defense against pathogens. They may also help regulate growth, differentiation, and flower development all by activating signal pathways.
Plant hormones, or plant growth regulators, are chemicals produced by plants that alter growth patterns. They can be found in many cells and tissues, although plant hormones seem to be concentrated in meristems and buds. One should not confuse hormones with enzymes. Any chemical reaction that occurs in a cell requires a specific catalyst. Enzymes are the catalysts found in cells. Hormones inhibit as well as promote cellular activities by sending chemical signals to cells.
The hormones identified in plants most often regulate division, elongation, and differentiation of cells. Plant hormones work in very small concentrations, affecting membrane properties, controlling gene expression, and affecting enzyme activity. In most cases, the effect plant hormones have on the plant depends on the location and concentration of the hormone relative to other hormones. Hormones often work in conjunction with each other, have overlapping effects, and work with environmental stimuli. There are several classes of plant hormones, including a number of recently “discovered” ones.
Auxins
The concept of chemical messengers in plants was proposed by Charles Darwin in 1881, who spent time looking at the phenomenon of phototropism in wheat seedlings. When a seedling is illuminated from the side, the shoot will bend towards the light. Darwin did a number of experiments and determined that a certain chemical traveled to the region of elongation and effected a differential elongation of cells furthest from the light sources. The chemical was subsequently studied and named auxin. Chemically, auxin is indoleacetic acid (IAA). A primary site of auxin production is the apical shoot meristem.
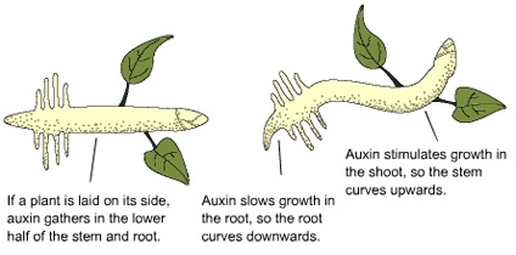{kind=link}
The most studied function of auxin is cell elongation when protons stimulate proteins that disrupt hydrogen bonds and break cross linkages in cellulose. This facilitates wall expansion when cells take in more water.
- Auxins migrate away from a light source.
- Auxin stimulates cambium cells to divide
- Auxin stimulates secondary xylem to differentiate.
- Auxin tends to inhibit the activation of buds lower on the stems. This is known as apical dominance.
- Cytokinins counter the apical dominance effect of auxins.
- Auxins promote lateral and adventitious root development.
- Auxins promote other hormone production.
- Auxins promote flower initiation.
- Loss of auxin initiates leaf abscission.
- Fruit development requires auxin produced by the developing seed.
- Auxins are toxic in large concentrations. They affect mostly dicots but not monocots. Herbicides made from synthetic auxins include 2-4-D and 2-4-5-T.
Cytokinins
Cytokinins are one of the molecules in DNA. Biologists have yet to identify the genes for the cytokinins found in plants. One hypothesis today is that cytokinins are actually made by symbiotic methyl bacteria that live within tree tissues. They are found primarily in root meristems, embryos, and fruits, and migrate from roots to the shoot systems of plants in xylem tissue.
The effects of cytokinins are often studied in tissue culture. Parenchyma cells grown in tissue culture will not divide and differentiate unless cytokinins and auxin are present. Cytokinins can promote auxiliary bud growth by overriding the inhibiting effect of auxin. This is one of the ways in which plants balance root and shoot growth.
Cytokinins also retard leaf death, by stimulating RNA and protein synthesis and delaying degradation of chlorophyll. Galls in plants take advantage of cytokinin function. The bacteria that form these tumors contain genes for the synthesis of cytokinins, which promote rapid cell growth.
Gibberellins
The discovery of gibberellins found that a fungus was responsible for abnormal growth. The fungus was Gibberella fujikuroi. Over 100 different gibberellins are known and are produced in roots and younger leaves. Gibberellins work with auxins to promote rapid elongation and division of stem tissue. This is seen in:
- reversal of genetic dwarfism,
- promotion of flowering in biennials during the first growing season, a process called bolting,
- gibberellins signaling germination activities,
- stimulation of some fruit enlargement,
- countering the effects of herbicides.
PBZ (Paclobutrazol)
PBZ is a gibberelin biosynthesis inhibitor that reduces the growth of many species and is commonly used on trees under utility lines or anywhere else the tree size needs to be controlled. The product has also been used to stimulate root regeneration after transplanting. It has been studied for the purpose of stabilizing declining trees that have insufficient fine root development. Treatment should be part of a complete tree care program including mulching.
Abscissic Acid
Abscissic acid (ABA) is a hormone that functions by inhibiting growth activities in times of environmental stress rather than by promoting growth. It got its name from the erroneous belief that it promoted the formation of abscission layers in leaves and fruits. It does not do this, although leaf abscission accompanies dormancy in many plants.
ABA promotes seed dormancy activities. ABA levels are high when seeds mature, promoting lowered metabolism and synthesis of proteins needed to withstand the dehydration associated with dormancy. Seeds germinate when ABA is degraded by some environmental incident such as when rain has washed the ABA out of the seed coat or light-stimulated enzymes. Breaking dormancy is relative to the ratio of ABA (which keeps seeds in dormancy) and gibberellins (which promote germination). Low levels of ABA in maturing seeds promotes premature germination.
ABA promotes stomata closure during leaf water deficit conditions. ABA in this case originates in roots, and detects low water level in root tissues, then moves upward and activates stomatal closure.
ABA derivatives, called dormins, are used in commercial nurseries to keep materials to be shipped in dormant conditions. The dormancy can be reversed with gibberellins.
Ethylene
Ethylene is a gas best known as a fruit ripener. Gardeners in China knew centuries ago that fruits ripened faster in rooms with burning incense. Citrus growers used kerosene stoves in the rooms in which they ripened their fruit. Today, grocer warehouses have ethylene rooms for ripening most of our produce. Ethylene affects many aspects of growth and development in tissues throughout the tree, but emphasis is on fruit maturation, leaf abscission, and senescence.
A shoot tip that encounters an immovable object will grow around the object by changing its growth direction. This occurs through differential elongation of cell walls as ethylene is synthesized, slowing cell wall expansion.
Brassinolides (Brassinosteroids)
Brassinolides is a tree steroid discovered in pollen of members of the mustard family, and is best studied in the Arabadopsis plant. Chemically they are very similar to animal steroid hormones. Brassinosteroids signal cell elongation and cell division. Brassinosteroids promote differentiation of xylem tissue. Brassinosteroids can also retard leaf abscission. Absence of brassinolides results in dwarf plants.
Salicylic Acid
Salicylic acid is known to activate defense genes against pathogen invaders. Salicylic acid, a phenolic extract from Salix (willow) bark, was long used as an analgesic. It is now prepared commercially and is the active ingredient of aspirin.
Jasmonates
Jasmonates are a group of fatty acid derivatives. They appear to have a role in seed germination, root growth, and the storage of protein in seeds. Synthesis of defense proteins may be triggered by jasmonates.
Systemin
Systemin is a small peptide found in wound tissue. It may stimulate defense activities in other parts of the tree to prevent more wounding.
Oligosaccharins
Oligosaccharins are short chain sugars in cell walls that may have a role in defense against pathogens. They may also help regulate growth, differentiation, and flower development all by activating signal pathways.
Failure of Roots
Up to 75% of all tree failures may be root related, and although most trees fail when winds exceed 50 mph, weak roots can fail anytime. Failure as defined here occurs when the roots fail to mechanically keep the tree standing. Being able to predict root failure is important and it is more difficult than predicting stem failure because the roots are hidden from view. When roots fail, the losses can be catastrophic.
Poor growth, thinning, chlorosis, and general decline manifest as root damage symptoms. Through a variety of injuries caused by construction, improper drainage, and soil compaction, roots can sustain damage. Crown symptoms mean that the root system has been compromised structurally, feeder roots have been affected, or the tree may have problems with its vascular system. These symptoms may warrant a root crown inspection. Visible signs of root damage include bleeding wounds on the trunk, loose bark around the flare, sunken areas on the trunk and flare, girdling roots and roots above the flare, and cracks extending into the stem from the soil line. Covering these root injuries with grass or soil will hide the damage, but not make the problem go away.
Three general types of failure associated with root systems have been identified.
1. Soil Failures – A common factor for soil failure is saturated soil and winds in excess of 50 miles per hour. This occurs after several days of heavy rain or excessive irrigation. Soil compaction, thin soil over rock ledge, hardpan layers, poor drainage areas, or high water tables can also cause failure and restrict root development. Poor soils fail to keep the tree securely anchored and account for 2/3 of the number of tree failures.
2. Root Failures – Root failures occur when the force of wind on the tree exceeds the strength of root wood. The roots that break are mainly 1 inch (2.5 cm) or smaller, regardless of the tree size. This occurs on healthy trees in winds that exceed 60 miles per hour, but is more common in trees with severely decayed or damaged roots. Soil covering the flare and girdling roots also increase the risk of failure. Root failures are recognized by the presence of large broken roots or wood fractures that begin at the roots and progress up the lower stem.
3. Human Factors – The most common failure of roots in urbanized areas are caused by root "pruning". This event occurs when roots have been cut by building and construction activities including the installation of sidewalks, curbs, and underground utilities. Regardless of whether the roots were pruned properly or ripped by construction equipment the results can be the same; tree failure occurs weeks to years after the damage was done. Proper pruning usually delays the tree failure but does not eliminate the potential for failure.
Root Decay
Root decay is difficult to recognize during a visual examination. Generally, healthy roots have intact bark and are whitish when the bark is removed. Rotting roots are usually blackish and have loose bark.
Decay typically begins at the root tips and progresses up the lower side of the root. The upper side of the root may not be affected, so most visual signs of root decay can not be easily seen. If decay is detected, assume that the remainder of the root below where the decay was found is likely compromised as well. This decay may also extend up the center of the lower trunk.
A recent study following a storm indicated that root rot was present in 13% of failed trees, while 38% of trees that survived the storm also contained root rot. Decay occurs most often in the roots of old trees that have been disturbed by construction some time in the past. Decay manifests its presence with fungal conks, bracts, or mushrooms. Inspectors should learn to recognize the fungal conks, bracts, or mushrooms that undermine root stability and grow in soil around the trunk or from the buttress roots. On conifers, cracks in the stem indicate root rot. Most arborists agree that trees that have lost more than 50% of their root systems should be considered at risk.
Tree Risk Assessment
The cutting of large buttress roots during construction can severely reduce the structural integrity of a tree. The minimum distance from the trunk where cuts should be made depends on the tree's condition and other factors listed below. Potential targets around the tree also need to be identified.
Tree and site assessments should determine the history of soil disturbance from construction and other causes. Tree exposure considerations include:
Hazard Determination
The point at which a tree should be considered hazardous or a managed risk is not always easy to determine. Root decay and other factors such as tree species, lean, and crown density must all be considered in their relationship to a target.
Thresholds have been determined for root decay and severance.
Thresholds should be reduced in cases of the tree exposure considerations indicated above, plus weak-wooded species, decayed roots that are opposite the lean (the tension-loaded roots), when the live crown ratio is less than 33%, or other risk factors when the tree owner is willing to accept only a slight risk.
Hazard Mitigation
Risk of root failure can be reduced in several ways.
When the roots and lower stem are visible they can be struck with a mallet to hear if they are hollow. Any roots that sound hollow should be probed to determine the thickness of solid wood with an increment borer, electric drill or decay-detecting drill. The amount of solid wood should be measured and recorded for each buttress root.
Root Injury May Cause Tree Failure Many Years Later
Large landscape trees can fall at any time. When this happens, it is often found that the tree failed because of events that damaged the tree's roots years earlier.
Along with other vital functions, roots hold a tree firmly in place. Anything that changes the structural support decreases the stability of the tree. A root system's ability to support the tree will decrease because of:
Harmful microorganisms that live in the soil often infect a root through a wound, but some are able to penetrate a root directly, especially if the tree has been weakened by drought or excess water. Some root rot fungi can kill a tree and it never falls; others cause living and healthy looking trees to fall. Sometimes trees blow over with the remains of their decayed root systems still attached.
It is usually impossible to predict the exact effect that root cutting will have on a particular tree, or when failure will actually occur. A tree may fail a few months or many years following root injury, despite the root injury. Tree species vary in their ability to tolerate root disturbances. A tree with a deep, extensive root system will tolerate more disturbance than a neighboring tree with a poorly developed root system. In general, it is important to take every step possible to avoid cutting or damaging a tree's root system.
Despite an occasional failure, most large trees are very safe. Root systems are well designed by nature to hold trees up, regardless of the tree's size. For the most part, they do just that.
Up to 75% of all tree failures may be root related, and although most trees fail when winds exceed 50 mph, weak roots can fail anytime. Failure as defined here occurs when the roots fail to mechanically keep the tree standing. Being able to predict root failure is important and it is more difficult than predicting stem failure because the roots are hidden from view. When roots fail, the losses can be catastrophic.
Poor growth, thinning, chlorosis, and general decline manifest as root damage symptoms. Through a variety of injuries caused by construction, improper drainage, and soil compaction, roots can sustain damage. Crown symptoms mean that the root system has been compromised structurally, feeder roots have been affected, or the tree may have problems with its vascular system. These symptoms may warrant a root crown inspection. Visible signs of root damage include bleeding wounds on the trunk, loose bark around the flare, sunken areas on the trunk and flare, girdling roots and roots above the flare, and cracks extending into the stem from the soil line. Covering these root injuries with grass or soil will hide the damage, but not make the problem go away.
Three general types of failure associated with root systems have been identified.
1. Soil Failures – A common factor for soil failure is saturated soil and winds in excess of 50 miles per hour. This occurs after several days of heavy rain or excessive irrigation. Soil compaction, thin soil over rock ledge, hardpan layers, poor drainage areas, or high water tables can also cause failure and restrict root development. Poor soils fail to keep the tree securely anchored and account for 2/3 of the number of tree failures.
{kind=link}
2. Root Failures – Root failures occur when the force of wind on the tree exceeds the strength of root wood. The roots that break are mainly 1 inch (2.5 cm) or smaller, regardless of the tree size. This occurs on healthy trees in winds that exceed 60 miles per hour, but is more common in trees with severely decayed or damaged roots. Soil covering the flare and girdling roots also increase the risk of failure. Root failures are recognized by the presence of large broken roots or wood fractures that begin at the roots and progress up the lower stem.
3. Human Factors – The most common failure of roots in urbanized areas are caused by root "pruning". This event occurs when roots have been cut by building and construction activities including the installation of sidewalks, curbs, and underground utilities. Regardless of whether the roots were pruned properly or ripped by construction equipment the results can be the same; tree failure occurs weeks to years after the damage was done. Proper pruning usually delays the tree failure but does not eliminate the potential for failure.
{kind=link}
Root Decay
Root decay is difficult to recognize during a visual examination. Generally, healthy roots have intact bark and are whitish when the bark is removed. Rotting roots are usually blackish and have loose bark.
Decay typically begins at the root tips and progresses up the lower side of the root. The upper side of the root may not be affected, so most visual signs of root decay can not be easily seen. If decay is detected, assume that the remainder of the root below where the decay was found is likely compromised as well. This decay may also extend up the center of the lower trunk.
A recent study following a storm indicated that root rot was present in 13% of failed trees, while 38% of trees that survived the storm also contained root rot. Decay occurs most often in the roots of old trees that have been disturbed by construction some time in the past. Decay manifests its presence with fungal conks, bracts, or mushrooms. Inspectors should learn to recognize the fungal conks, bracts, or mushrooms that undermine root stability and grow in soil around the trunk or from the buttress roots. On conifers, cracks in the stem indicate root rot. Most arborists agree that trees that have lost more than 50% of their root systems should be considered at risk.
Tree Risk Assessment
The cutting of large buttress roots during construction can severely reduce the structural integrity of a tree. The minimum distance from the trunk where cuts should be made depends on the tree's condition and other factors listed below. Potential targets around the tree also need to be identified.
Tree and site assessments should determine the history of soil disturbance from construction and other causes. Tree exposure considerations include:
- What is the height and spread of the crown?
- What factors of the location are significant, including wind direction?
- Does the tree lack branches on the lower 2/3 of the stem (live crown ratio less than 33%)?
- Is it growing on a slope?
- What is the condition of the roots?
- Any unusual swelling or depression at the base of the tree?
- Are the buttress roots visible and undamaged?
- Are any signs of decay fungi present?
- Has there been recent construction near the tree, including fill over the flare?
- Are there girdling roots?
- What are the soil types, depth, and width?
- What is the depth to ground water and bedrock?
Hazard Determination
The point at which a tree should be considered hazardous or a managed risk is not always easy to determine. Root decay and other factors such as tree species, lean, and crown density must all be considered in their relationship to a target.
Thresholds have been determined for root decay and severance.
- Major buttress roots are considered significantly decayed if the thickness of sound wood on the root is less than the tree diameter (DBH) times 0.15. So, on a tree 30 inches (57.5 cm) dbh, each major root must have at least 4½ inches (11 cm) of solid wood. If it has less, it is considered severely decayed.
- Major buttress roots that were cut within a distance of three times the trunk diameter are considered a high risk.
- If more than a third of the major buttress roots are missing or decayed, the tree should be considered a
high risk - If more than half of the major buttress roots are missing or decayed, the tree should be considered a critical risk.
Thresholds should be reduced in cases of the tree exposure considerations indicated above, plus weak-wooded species, decayed roots that are opposite the lean (the tension-loaded roots), when the live crown ratio is less than 33%, or other risk factors when the tree owner is willing to accept only a slight risk.
Hazard Mitigation
Risk of root failure can be reduced in several ways.
- If possible, move the target away to a distance of 1½ times the tree height. This technique may require installation of fencing around the tree to keep people out of the target area.
- If the risk is low or moderate, pruning to reduce the percentage of crown may lower the wind resistance.
- For high and critical risk trees, the tree should be removed.
When the roots and lower stem are visible they can be struck with a mallet to hear if they are hollow. Any roots that sound hollow should be probed to determine the thickness of solid wood with an increment borer, electric drill or decay-detecting drill. The amount of solid wood should be measured and recorded for each buttress root.
Root Injury May Cause Tree Failure Many Years Later
Large landscape trees can fall at any time. When this happens, it is often found that the tree failed because of events that damaged the tree's roots years earlier.
Along with other vital functions, roots hold a tree firmly in place. Anything that changes the structural support decreases the stability of the tree. A root system's ability to support the tree will decrease because of:
- root decay or rots,
- cutting large roots near the base of the tree,
- drought,
- flooding or over-watering,
- grade changes,
- soil compaction,
- gas leaks,
- paving over roots.
Harmful microorganisms that live in the soil often infect a root through a wound, but some are able to penetrate a root directly, especially if the tree has been weakened by drought or excess water. Some root rot fungi can kill a tree and it never falls; others cause living and healthy looking trees to fall. Sometimes trees blow over with the remains of their decayed root systems still attached.
It is usually impossible to predict the exact effect that root cutting will have on a particular tree, or when failure will actually occur. A tree may fail a few months or many years following root injury, despite the root injury. Tree species vary in their ability to tolerate root disturbances. A tree with a deep, extensive root system will tolerate more disturbance than a neighboring tree with a poorly developed root system. In general, it is important to take every step possible to avoid cutting or damaging a tree's root system.
Despite an occasional failure, most large trees are very safe. Root systems are well designed by nature to hold trees up, regardless of the tree's size. For the most part, they do just that.
Root Failure by Fungi
Vigorously growing trees are usually able to resist decay fungi by outgrowing them or by stopping them with natural barriers, but wounded or weakened trees are more susceptible to invasion. Some of the fungi that cause serious tree diseases are easily controlled by fungicide applications. Unfortunately, the fungicide will also kill beneficial fungi. Many older trees that fail during storms have had their strength reduced by decay fungi. Strength loss is difficult to detect but mushrooms or bracket fungi on the trunk are warnings. Even after the tree has died, these fungi continue to use the tree roots as a food base, which they can transfer to new trees by root contact or by spores released from fruiting bodies (conks).
Armillaria
Armillaria root disease, also known as shoestring root rot, is caused by the fungus Armillaria. Forty species of Armillaria cause death to trees under all types of stress by reducing absorption and translocation of water and minerals from the soil to the tree. However, because many symptoms produced by Armillaria-infected trees are nonspecific, they are easily misdiagnosed as drought, mineral deficiencies, infestations, and other pathogen attacks. Confirmation of Armillaria root disease is based on finding toadstools that form between the bark and wood of buttress roots; black rhizomorphs (shoestring-like aggregations of mycelium) on infected roots, under the bark of infected trees, and in the soil near the infected trees. The fungus survives and spreads in organic mulches. Other Armillaria include:
Management Recommendations
Annosus Root Disease
Heterobasidion annosum (formerly called Fomes annosus) is found in temperate zone forests, where it causes butt rot of conifers. The P-type is found only on pines and the S-type is found on spruce and other conifers. The P-type occurs from Eastern North America to the Rocky Mountains. The west coast has both S and P-types and these are regarded as serious problems. The fungus infects freshly cut stumps by means of airborne spores, growing into its roots, and spreading to adjacent healthy trees at root contacts. Airborne spores also infect stem wounds on thin-barked species, such as hemlock and fir, where extensive decay can develop above and below the wound. It forms conks. Pines tend to be killed outright when the fungus grows in the cambium and girdles the trunk at the flare. Other conifers tend to get butt rot but can survive for many years with the disease.
Another related fungus is Fomes fomentarius which infects maple, beech, and birch, commonly on older pruning wounds.
Stumps in the West may sustain the fungus for over 50 years while in the Southeast, the fungus can consume a stump in less than 10 years.
Management Recommendations
Laminated Root Rot (LRR)
There are three forms of the fungus:
1. Phellinus weirii: "Asian" form,
2. "North American Douglas-fir" form known as P. sulphurascens,
3. "Cedar" form.
The Douglas-fir form affects several conifer species while the Cedar form affects species of cedar. This is one of the most damaging root diseases affecting conifers in northwestern North America. All hardwoods are immune to LRR infection. The disease causes mortality, growth reduction, and butt rot. LRR first infects and kills its host, then colonizes and uses them as food sources. LRR begins when roots contact infected stumps from the previous stand. LRR may survive in stumps for up to 50 years and will produce brown, flat conks. LRR can colonize healthy root bark, although older trees are better able to tolerate infection.
Management Recommendations
White Pine Root Disease
Verticicladiella procera is of great significance in Christmas tree plantations and in new forest plantations. It is also associated with dying white pine stands in the southern Appalachians. There are no fruiting bodies associated with this fungus that can be readily seen. Infected, mature white pines die from the top down. Some trees may die within a year and others may decline over several years, with mortality randomly occurring, with 1% to 3% of the affected trees dying annually. A chocolate-brown canker may occur under the bark at the flare. Tree death may result from the killing of numerous small roots 3/16 inch (5 mm) in diameter or less. Wet sites appear to favor the disease.
Management Recommendations
Other Fungi
Root and butt rots – These are wood-decay diseases caused by Basidiomycota. They enter through wounds in the trunk or roots and kill cambial tissues. The tree will die when the flare is girdled. A tree can live with butt rot and stem decay for many years. The tree may often die because it is uprooted or snapped, rather than from the rot itself.
Cerrena unicolor – fruits as multiple, thin, leathery shelves, with green moss on top of younger shelves. It
produces an ever-expanding elongated canker on many deciduous trees.
Climacodon septentrionalis – is made up of multiple cream-colored, spore bearing shelves that fruits in
August and September, especially on sugar maple.
Cortical root rots – Many of the fungi that attack the cortex of young roots move into the cambium and phloem of woody roots.
Daedalea quercina – has woody, shelf-like fruiting bodies growing from old pruning wounds. The underside
is maze-like and this fungus is common on oaks.
Artist conk – Ganoderma applanatum is a woody shelf-type fruiting body up to 2 ft (60 cm) wide that is found on the lower part of trunk. The top is brownish with a creamy white margin. Its wide host range includes oaks, maples, and beech.
Chicken-of-the-woods – Laetiporus sulfureus has a lemon-yellow/orange shelf-type fruiting body found singly or in overlapping fans on trunks and butts of many deciduous trees.
Black-Stain Root Disease – Various western conifers will reveal dramatic streaks of black stain caused by Leptographium wageneri, a deuteromycete that is closely related to Ophiostoma stain fungi. The three hosts are Ponderosa pine, Pinon pine, and Douglas-fir. Root feeding weevils are attracted to fresh wounds and stumps and thinning sometimes initiates the disease. The dark hyphae grow in tracheids and are probably insect-vectored for long-distance dispersal. It grows root-to-root for secondary spread. Avoid wounding, creating wet areas, and performing commercial thinning, and conduct operations in summer to avoid vector activity.
Tomentosus Root Disease – Inonotus tomentosus is known to be the most widespread root disease in the boreal and sub-boreal forests of the northern hemisphere. The host range includes spruce and larch. The disease causes mortality, growth reduction, root infections, butt rot, and predisposition to wind-throw. The effects of Tomentosus vary with geographic region, soil factors, pathogen spread, and host response. This fungus can build up in the stumps with continued cutting. Spores can spread the disease quickly to new areas. I. tomentosus can survive in stumps for up to 30 years. Research has led to control by host resistance and removal of infected stumps. Commercial thinning of susceptible species can encourage planting and establishment of more resistant species.
Rhizoctonia Root Rot – Rhizoctonia solani is a root rot and web blight that can infect all ornamental plants. It spreads most quickly in warm, wet, and humid conditions. It grows within the top several inches of the soil and will spread from infected roots into the lower branches, killing leaves and stems very quickly.
Black Root Rot – This disease is caused by Thielaviopsis basicola and can be devastating. It is usually found in palms, Japanese holly, and Illicium, as well as phlox, verbena, and hibiscus. Leaves will turn yellow shortly before death. The roots become black and water soaked. The disease is more severe when the temperature is high and the pH is over 6.0. Sanitation is the best control and fungicides are useful when the disease is detected.
Phytophthora fungi – This is a major problem in many areas of the United States and other countries as well.
Editor's note: The use of trade names in this article is meant for the convenience of the reader and does not constitute an endorsement or approval of any product to the exclusion of others.
Sources
The test that follows contains 80 questions. Before taking the test be sure you have read the article carefully. The passing grade is 80% on the entire test.
ISA will award 4.0 CEUs* for a passing grade. SAF members will earn 1.0 Cat. 1-CT credit for a passing grade. The cost for taking this test is $20 per credit. If you purchase an annual subscription for 15 credits, the cost per credit is reduced by 50% (see Annual Subscription link below). We will report all passing test scores to ISA and/or SAF. If you are a member of ISA and SAF we will report your passing test scores to both for no additional cost. Tests with passing scores may be submitted only once to each organization.
*Members of ISA may apply the 4.0 CEUs toward Certified Arborist, Tree Worker Specialist, Municipal Specialist, or BCMA science credits.
California UFC members will receive credit for passing the test. Please add your CaUFC number after your ISA and/or SAF certification number.
ASCA and MTOA members may submit your ISA certification record to these organizations and receive credits one for one.
To take the test by the pay per test option, click on the 'Pay Now' button below where you can send payment online securely with your credit card or Pay Pal account. After your payment is submitted, click on ‘Return to Merchant' / gibneyCE.com. That will take you to the test sign in page followed by the test. Members with certifications from both ISA and SAF, please be sure to add both of your certification numbers. These numbers are important for reporting purposes.
To take the test as an annual subscriber with reduced rates, click on Password and enter your test password which will take you to the test sign in page. If you would like to become a subscriber see our Annual Subscription page for details.
When you have finished answering all questions you will be prompted to click ‘next’ to send your answers to gibneyCE.com. You can then click ‘next’ to view your test summary. A test review of your answers is available upon request.
All passing test scores are sent from gibneyCE.com to your organization(s) at the end of every month and they will appear on your certification record 4 to 6 weeks* after that. ISA maintains a record of CEU credits on their website *SAF requires 5 passing test scores before reporting.
Test re-takes are allowed, however you will have to pay for the retake if you are using the pay per test option. You can spend as much time as you would like to take the test but it is important not to leave the test site until you have answered all the questions and see the 'sending your answers' response.
Vigorously growing trees are usually able to resist decay fungi by outgrowing them or by stopping them with natural barriers, but wounded or weakened trees are more susceptible to invasion. Some of the fungi that cause serious tree diseases are easily controlled by fungicide applications. Unfortunately, the fungicide will also kill beneficial fungi. Many older trees that fail during storms have had their strength reduced by decay fungi. Strength loss is difficult to detect but mushrooms or bracket fungi on the trunk are warnings. Even after the tree has died, these fungi continue to use the tree roots as a food base, which they can transfer to new trees by root contact or by spores released from fruiting bodies (conks).
Armillaria
Armillaria root disease, also known as shoestring root rot, is caused by the fungus Armillaria. Forty species of Armillaria cause death to trees under all types of stress by reducing absorption and translocation of water and minerals from the soil to the tree. However, because many symptoms produced by Armillaria-infected trees are nonspecific, they are easily misdiagnosed as drought, mineral deficiencies, infestations, and other pathogen attacks. Confirmation of Armillaria root disease is based on finding toadstools that form between the bark and wood of buttress roots; black rhizomorphs (shoestring-like aggregations of mycelium) on infected roots, under the bark of infected trees, and in the soil near the infected trees. The fungus survives and spreads in organic mulches. Other Armillaria include:
- Armillaria sinapina is the most common species and it kills broadleaf trees and shrubs. A. sinapina often colonizes on conifer stumps in managed coastal forests.
- A. gallica spreads slowly on the host's root system; usually only stressed hosts are killed.
- A. ostoyae causes significant damage, including growth repression and mortality to conifers, broadleaf trees, shrubs, and some herbs. It is capable of penetrating, infecting, and killing healthy, vigorous trees. Entry into host trees occurs through roots, wounds, and directly through uninjured tissues.
Management Recommendations
- Reduce environmental stress and encourage and maintain tree vigor.
- Maintain the natural composition of tree species.
- Replant with resistant or moderately resistant species.
- Use mycorrhizae as a non-chemical control of Armillaria.
- Harvest conventionally and remove the stump to reduce inoculum.
- Prevent spread of rhizomorphs by trenching around infected trees.
- Remove dead and dying trees during logging operations.
- Avoid wounding residual trees during logging operations.
- Plow and burn stumps of Armillaria-infected trees before replanting.
- Delay planting on land previously occupied by oaks.
Annosus Root Disease
Heterobasidion annosum (formerly called Fomes annosus) is found in temperate zone forests, where it causes butt rot of conifers. The P-type is found only on pines and the S-type is found on spruce and other conifers. The P-type occurs from Eastern North America to the Rocky Mountains. The west coast has both S and P-types and these are regarded as serious problems. The fungus infects freshly cut stumps by means of airborne spores, growing into its roots, and spreading to adjacent healthy trees at root contacts. Airborne spores also infect stem wounds on thin-barked species, such as hemlock and fir, where extensive decay can develop above and below the wound. It forms conks. Pines tend to be killed outright when the fungus grows in the cambium and girdles the trunk at the flare. Other conifers tend to get butt rot but can survive for many years with the disease.
Another related fungus is Fomes fomentarius which infects maple, beech, and birch, commonly on older pruning wounds.
Stumps in the West may sustain the fungus for over 50 years while in the Southeast, the fungus can consume a stump in less than 10 years.
Management Recommendations
- Reduce wounding in all trees especially conifers (except pines), and treat stumps with chemicals (borax) or biocontrol (Phlebia gigantea).
- Plant and thin during times when infection risk is minimal, from May to August.
- Replace susceptible hosts with resistant species and encourage mixed-species stands.
Laminated Root Rot (LRR)
There are three forms of the fungus:
1. Phellinus weirii: "Asian" form,
2. "North American Douglas-fir" form known as P. sulphurascens,
3. "Cedar" form.
The Douglas-fir form affects several conifer species while the Cedar form affects species of cedar. This is one of the most damaging root diseases affecting conifers in northwestern North America. All hardwoods are immune to LRR infection. The disease causes mortality, growth reduction, and butt rot. LRR first infects and kills its host, then colonizes and uses them as food sources. LRR begins when roots contact infected stumps from the previous stand. LRR may survive in stumps for up to 50 years and will produce brown, flat conks. LRR can colonize healthy root bark, although older trees are better able to tolerate infection.
Management Recommendations
- In sapling stands, cull all trees that are susceptible to LRR.
- Remove infected trees completely so the fungus will die out.
- Salvage symptomatic trees and consider harvesting earlier than usual.
- Thinning is ineffective because there will be infected trees left as crop trees.
- When planting, an alternating mixture of species should be considered.
- Use resistant trees.
- Consider biological controls such as Tricoderma viride as soon as a suitable field delivery system is developed.
White Pine Root Disease
Verticicladiella procera is of great significance in Christmas tree plantations and in new forest plantations. It is also associated with dying white pine stands in the southern Appalachians. There are no fruiting bodies associated with this fungus that can be readily seen. Infected, mature white pines die from the top down. Some trees may die within a year and others may decline over several years, with mortality randomly occurring, with 1% to 3% of the affected trees dying annually. A chocolate-brown canker may occur under the bark at the flare. Tree death may result from the killing of numerous small roots 3/16 inch (5 mm) in diameter or less. Wet sites appear to favor the disease.
Management Recommendations
- Prevention is the best defense.
- Promote reasonable growth of young trees.
- Water well during dry spells.
- Aerate the soil.
- Maintain a 2" layer of organic mulch.
- Apply slow-release fertilizer in early autumn or spring.
- Efforts should be made to avoid injuring the roots during construction.
- Protect the young trunk from injury.
- Use proper pruning techniques, timing, and management.
Other Fungi
Root and butt rots – These are wood-decay diseases caused by Basidiomycota. They enter through wounds in the trunk or roots and kill cambial tissues. The tree will die when the flare is girdled. A tree can live with butt rot and stem decay for many years. The tree may often die because it is uprooted or snapped, rather than from the rot itself.
{kind=link}
Cerrena unicolor – fruits as multiple, thin, leathery shelves, with green moss on top of younger shelves. It
produces an ever-expanding elongated canker on many deciduous trees.
Climacodon septentrionalis – is made up of multiple cream-colored, spore bearing shelves that fruits in
August and September, especially on sugar maple.
Cortical root rots – Many of the fungi that attack the cortex of young roots move into the cambium and phloem of woody roots.
Daedalea quercina – has woody, shelf-like fruiting bodies growing from old pruning wounds. The underside
is maze-like and this fungus is common on oaks.
Artist conk – Ganoderma applanatum is a woody shelf-type fruiting body up to 2 ft (60 cm) wide that is found on the lower part of trunk. The top is brownish with a creamy white margin. Its wide host range includes oaks, maples, and beech.
Chicken-of-the-woods – Laetiporus sulfureus has a lemon-yellow/orange shelf-type fruiting body found singly or in overlapping fans on trunks and butts of many deciduous trees.
Black-Stain Root Disease – Various western conifers will reveal dramatic streaks of black stain caused by Leptographium wageneri, a deuteromycete that is closely related to Ophiostoma stain fungi. The three hosts are Ponderosa pine, Pinon pine, and Douglas-fir. Root feeding weevils are attracted to fresh wounds and stumps and thinning sometimes initiates the disease. The dark hyphae grow in tracheids and are probably insect-vectored for long-distance dispersal. It grows root-to-root for secondary spread. Avoid wounding, creating wet areas, and performing commercial thinning, and conduct operations in summer to avoid vector activity.
Tomentosus Root Disease – Inonotus tomentosus is known to be the most widespread root disease in the boreal and sub-boreal forests of the northern hemisphere. The host range includes spruce and larch. The disease causes mortality, growth reduction, root infections, butt rot, and predisposition to wind-throw. The effects of Tomentosus vary with geographic region, soil factors, pathogen spread, and host response. This fungus can build up in the stumps with continued cutting. Spores can spread the disease quickly to new areas. I. tomentosus can survive in stumps for up to 30 years. Research has led to control by host resistance and removal of infected stumps. Commercial thinning of susceptible species can encourage planting and establishment of more resistant species.
Rhizoctonia Root Rot – Rhizoctonia solani is a root rot and web blight that can infect all ornamental plants. It spreads most quickly in warm, wet, and humid conditions. It grows within the top several inches of the soil and will spread from infected roots into the lower branches, killing leaves and stems very quickly.
Black Root Rot – This disease is caused by Thielaviopsis basicola and can be devastating. It is usually found in palms, Japanese holly, and Illicium, as well as phlox, verbena, and hibiscus. Leaves will turn yellow shortly before death. The roots become black and water soaked. The disease is more severe when the temperature is high and the pH is over 6.0. Sanitation is the best control and fungicides are useful when the disease is detected.
Phytophthora fungi – This is a major problem in many areas of the United States and other countries as well.
Editor's note: The use of trade names in this article is meant for the convenience of the reader and does not constitute an endorsement or approval of any product to the exclusion of others.
Sources
- Ammon, Vernon, "Ornamental & Tree Diseases", Plant Disease Dispatch Sheets, Mississippi State University, December 2000.
- Appleton, Bonnie Lee, "Mycorrhizae", City Trees, Vol 37, Number 6 November/December 2001.
- Appleton, Bonnie, et.al., "Mycorrhizal Inoculation of Street Trees", Journal of Arboriculture Vol. 29, No. 2.
- Buschena, Cindy, "Will Mycorrhizal Inoculations Save Your Ailing Tree", Minnesota, ADVOCATE, Summer 2000.
- Carlson, Julie, et al, "Can Mycorrhizae Improve Tree Establishment in the Landscape?", SNA NewsLine, November/December, 2000.
- CFS research staff, "Tomentosus Root Disease", Canadian Forestry Service, 2008.
- Coder, Dr. Kim D., "Tree Root Growth Requirements", City Trees, The Journal of The Society of
Municipal Arborists, Vol. 38, Number 2, March/April 2002. - Goff, Kate, "Life Underground", Erosion Control, September/October 1999.
- "Guide to Arbor Care", Plant Health Care, Inc. 2011.
- Hart, Miranda M. and Richard J. Reader, "Variation In Arbuscular Mycorrhizae Fungi" New Phytology 2002.
- "Insects and Diseases of Trees in the South", USDA Forest Service - Southern Region, Protection Report R8-PR 16, June 1989.
- Johnson, David W., "Introduction to Decays", Forest Health Management, USDA Forest Service, Rocky Mountain Region, 1998.
- Kane, Brian & H. Dennis P. Ryan, "Tree Roots and Hazard Tree Evaluation", TCI Magazine, December 2002.
- Kernan, Michael J. PhD, "Biological Treatments for City Trees", City Trees, Vol 40, Number 3 May/June
2004. - Keslick and Son, "Technical Tree Biology Dictionary", Modern Arboriculture Associates, 2003.
- Little, Jane Braxton,"Mycorrhizae: Unsung Heroes", California Trees, Winter 2003 Vol. 14 No. 4.
- Mulrean Ed, PhD, "Root and Soil Failure", Arid Zone Trees, 2005.
- Neely, Dan, and Gary Watson, "The Landscape Below Ground II", International Society of Arboriculture,
1998. - Perry, Ed, "Root Injury May Cause Tree Failure Many Years Later", University of California Cooperative Extension, 1999.
- Phillips, Leonard, "Root Physiology", City Trees, The Journal of The Society of Municipal Arborists
Vol. 35, Number 4, July/August 1999. - Richardson, Rosemary, "Biology 203 Course", Bellevue Community College Science Division, Spring 2009.
- Rindels, Sherry, "Tree Root Systems", ISU Extension, Prepared by Department of Horticulture, Iowa State University, Ames, Iowa, April 1, 1992.
- "RHS research projects: plant pests & diseases", The Royal Horticultural Society 2011.
- Sinclair, Wayne A., Howard H. Lyon, and Warren T. Johnson, Diseases of Trees and Shrubs, Cornell University Press, Ithaca NY, 1987.
- Smiley, Thomas, PhD, "The Failure Potential of Roots", Shade Tree Technical Reports, Bartlett Tree Research Laboratories, 2010.
- Spafford, Anne, "Mycorrhizae", Landscape Architecture, February, 2001.
- Stabler, Linda B. et al., "Urbanization Effects on Tree Growth", Journal of Arboriculture 27(4):
193-202. - Topa, Mary A. PhD, "Root Physiology", The Encyclopedia of Forest Sciences, 2007.
- "Transplanting and Root Growth", City Trees, Vol 38, Number 1 January/February 2002.
- Watson, Gary W. and E. B. Himelick, "Principals and Practice of Planting Trees and Shrubs"', International Society of Arboriculture, Savoy, IL 1997.
- Wohlleben, Peter, "The Hidden Live of Trees", Greystone Books, 2016.
- William-Woodward, Jean, "How to Prevent Root Rot", Nursery Management and Production, September 2004.
- Worrall, J.J., "Forest & Shade Tree Pathology", Online Textbook, 2011.
The test that follows contains 80 questions. Before taking the test be sure you have read the article carefully. The passing grade is 80% on the entire test.
ISA will award 4.0 CEUs* for a passing grade. SAF members will earn 1.0 Cat. 1-CT credit for a passing grade. The cost for taking this test is $20 per credit. If you purchase an annual subscription for 15 credits, the cost per credit is reduced by 50% (see Annual Subscription link below). We will report all passing test scores to ISA and/or SAF. If you are a member of ISA and SAF we will report your passing test scores to both for no additional cost. Tests with passing scores may be submitted only once to each organization.
*Members of ISA may apply the 4.0 CEUs toward Certified Arborist, Tree Worker Specialist, Municipal Specialist, or BCMA science credits.
California UFC members will receive credit for passing the test. Please add your CaUFC number after your ISA and/or SAF certification number.
ASCA and MTOA members may submit your ISA certification record to these organizations and receive credits one for one.
To take the test by the pay per test option, click on the 'Pay Now' button below where you can send payment online securely with your credit card or Pay Pal account. After your payment is submitted, click on ‘Return to Merchant' / gibneyCE.com. That will take you to the test sign in page followed by the test. Members with certifications from both ISA and SAF, please be sure to add both of your certification numbers. These numbers are important for reporting purposes.
To take the test as an annual subscriber with reduced rates, click on Password and enter your test password which will take you to the test sign in page. If you would like to become a subscriber see our Annual Subscription page for details.
When you have finished answering all questions you will be prompted to click ‘next’ to send your answers to gibneyCE.com. You can then click ‘next’ to view your test summary. A test review of your answers is available upon request.
All passing test scores are sent from gibneyCE.com to your organization(s) at the end of every month and they will appear on your certification record 4 to 6 weeks* after that. ISA maintains a record of CEU credits on their website *SAF requires 5 passing test scores before reporting.
Test re-takes are allowed, however you will have to pay for the retake if you are using the pay per test option. You can spend as much time as you would like to take the test but it is important not to leave the test site until you have answered all the questions and see the 'sending your answers' response.